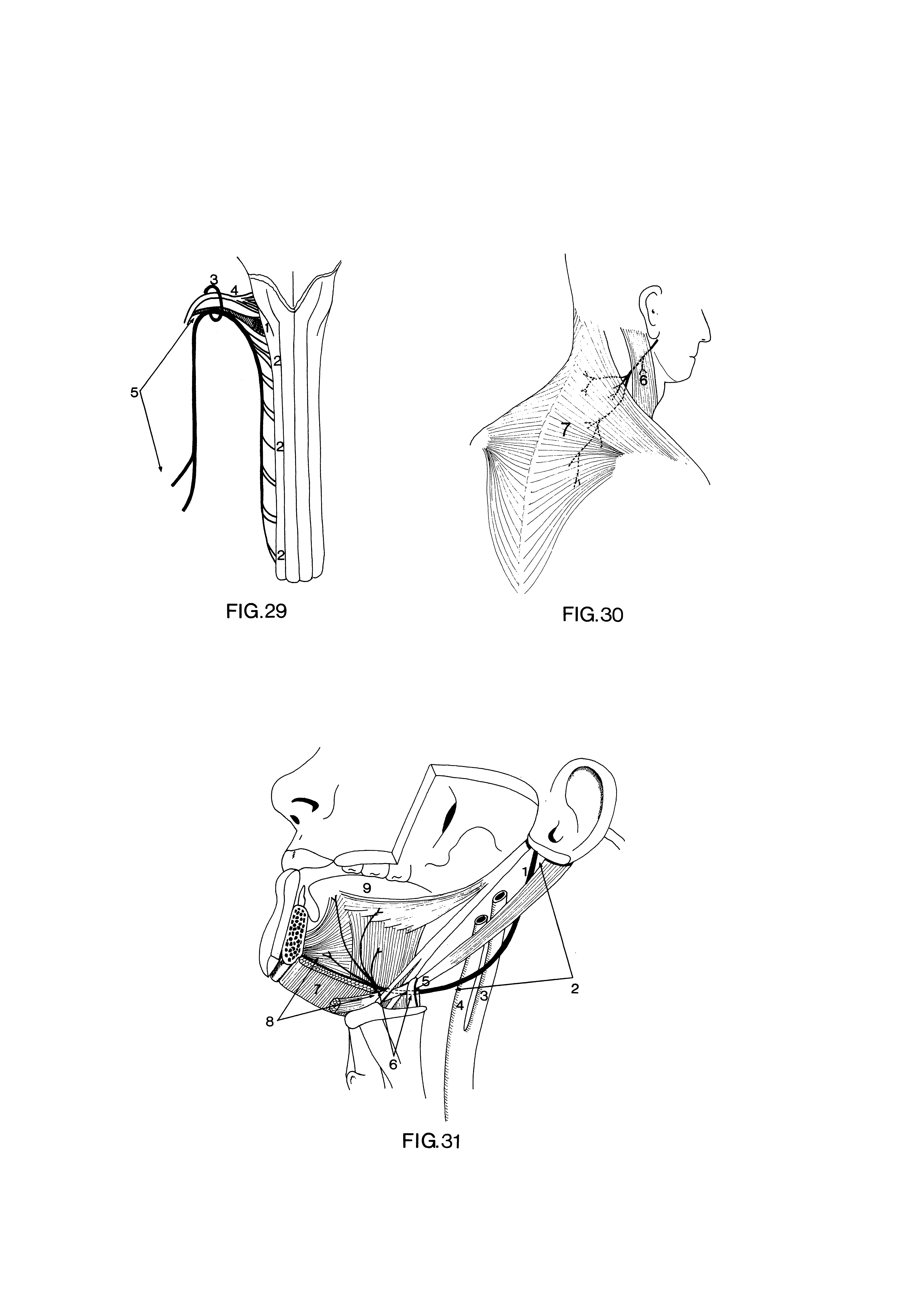
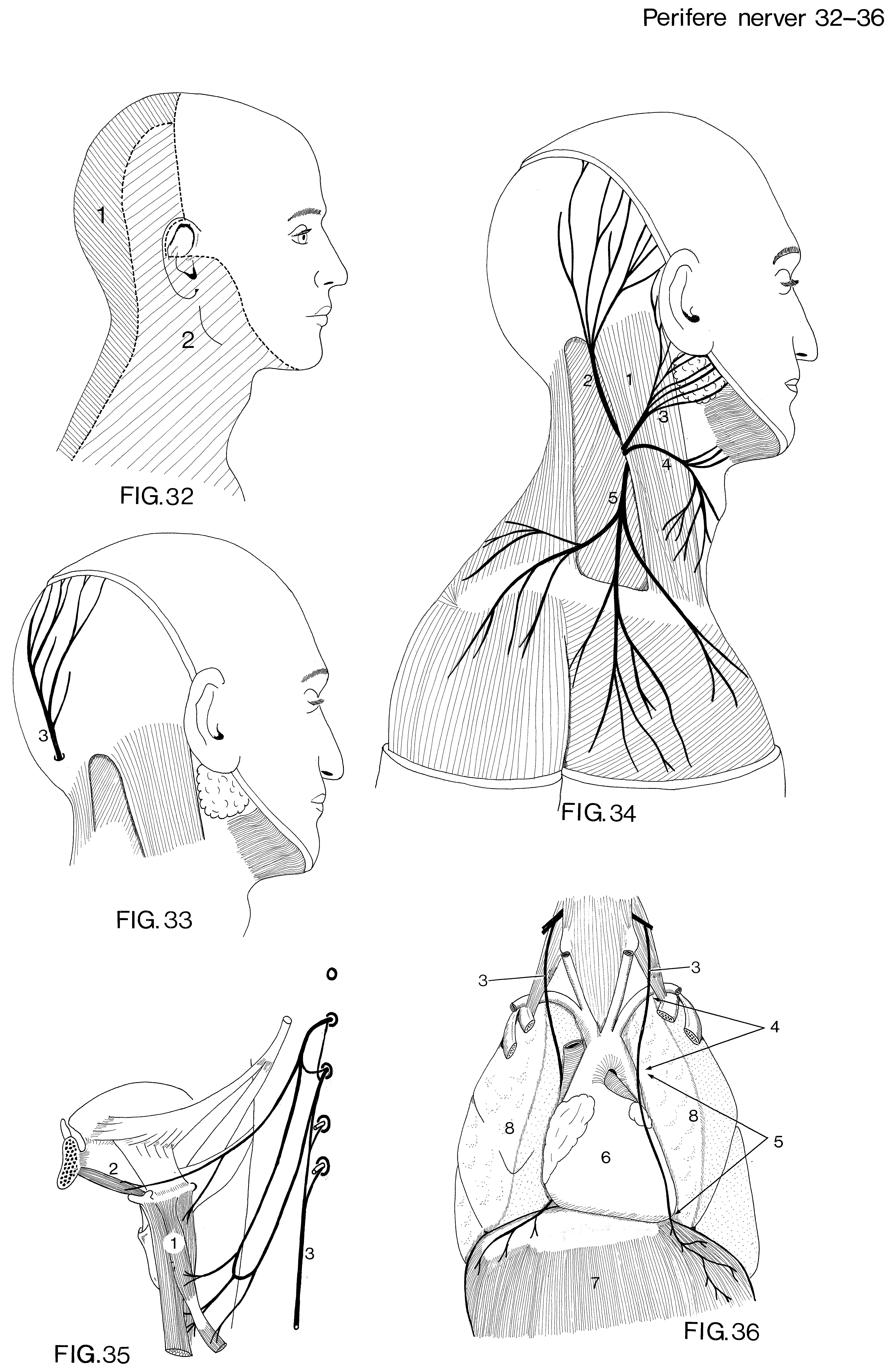
| Nervesystemets vigtigste grundelement er nervecellen, som er specialiseret i hurtig ledning af signaler over lange afstande. Tilsammen danner milliarder af nerveceller komplicerede og velordnede netværk i hjernen med henblik på kommunikation og informationsbehandling. | |
| Nervesystemet er udstyret med sanseorganer, receptorer, der opfanger sanseinformation = stimuli. Uanset typen af stimulus vil sanseorganet, receptoren, “oversætte” stimulusenergien til det “sprog”, nervesystemet forstår, nemlig nerveimpulser, der er små elektriske udladninger, som forplanter sig hurtigt langs nervetrådene. På den måde ledes signalerne fra receptorerne hurtigt til de dele af nervesystemet, hvor informationsbehandlingen foregår. | |
| Når nervesystemet skal frembringe en reaktion på en stimulus fra en receptor, sker det ved påvirkning af en effektor, som kan være enten en muskelcelle eller en kirtelcelle. Det betyder, at svaret sker i form af enten bevægelse eller sekretion. Sekretet kan have metaboliske virkninger, eller selv være et signal til forandring i andre celler. Muskelkontraktion kan vise sig på mange måder, lige fra kommunikation gennem tale, mimik og kropsattitude til flytning/bevægelse af kroppen, åndedræt og ændring af blodtryk. For at udtrykke sin “vilje”, har nervesystemet kun muligheden for påvirkning af muskelceller og kirtelceller. |
| På rent anatomisk grundlag inddeles nervesystemet i et centralnervesystem, der udgøres af rygmarv, medulla spinalis, og hjerne, encephalon – og det perifere nervesystem, som udgøres af de nerver, der sørger for kommunikationen mellem centralnervesystem og receptorer eller effektorer. |
Side 295
| Nervesystemet kan også betragtes som to dele uden helt så skarpe adskillelser, som den rent anatomiske inddeling. Den ene del regulerer først og fremmest indvoldsorganerne og kroppens indre miljø. Den anden del styrer vores tilpasning til omgivelserne. Den første del kaldes det autonome eller vegetative nervesystem. Da denne del hovedsageligt er knyttet til indvoldsorganer, viscera, benævnes det også det viscerale nervesystem.Den anden del kaldes gerne det somatiske eller cerebrospinale nervesystem. | |
| Betegnelsen autonom refererer til, at det stort set fungerer selvstændigt uden at være underkastet viljens herredømme eller kommer til individets bevidsthed. Dette i modsætning til det somatiske system, som i langt højere grad står under bevidsthedens indflydelse og kontrol. | |
| Den vigtigste opgave for det autonome nervesystem er at bidrage til opretholdelse af et stabilt indre miljø i legemet, = homeostasis, fx regulation af åndedræt, blodtryk, legemstemperatur og fordøjelse. Svigt i en homeostatisk mekanisme er ofte grundlaget for en sygelig proces.Det autonome nervesystem fungerer ikke altid uden deltagelse af det somatiske. Mange livsytringer er et produkt af et samspil mellem de to nervesystemer. Eksempelvis reagerer organismen på en kuldepåvirkning med sammentrækning af hudens blodkar, således at varmetabet mindskes. Det skyldes en refleks, hvor impulser fra kuldereceptorer ledes centralt gennem somatiske baner, mens karsammentrækkende autonome impulser ledes perifert. | |
| Det autonome nervesystem arbejder med lavere hastighed end det somatiske, idet de autonome nervetråde er tynde med lav ledningshastighed i modsætning til de stærkt myeliniserede somatiske nervetråde. En interessant forskel mellem de to systemer er også, at effektorcellen i det autonome nervesystem, fx en glat muskelcelle eller en kirtelcelle, kan fungere uden at atrofiere, selvom innervationen mistes. Afbrydes nerveforsyningen i det somatiske nervesystem til en tværstribet muskeltråd, vil den atrofiere og ophøre med at fungere. |
| Fælles for rygmarv og hjerne er, at de omgives af tre hinder og er godt beskyttet af knogle. Medulla spinalis ligger i canalis vertebralis og encephalon i cavitas cranii. |
| 12 par hjernenerver, nervi craniales, og 31 par rygmarvsnerver, nervi spinales. Hjernenerver og rygmarvsnerver kan under ét kaldes cerebrospinale nerver. |
Side 296
| Centralnervesystemets inddeling forstås bedre, når hovedtrækkene af udviklingen kendes. | |
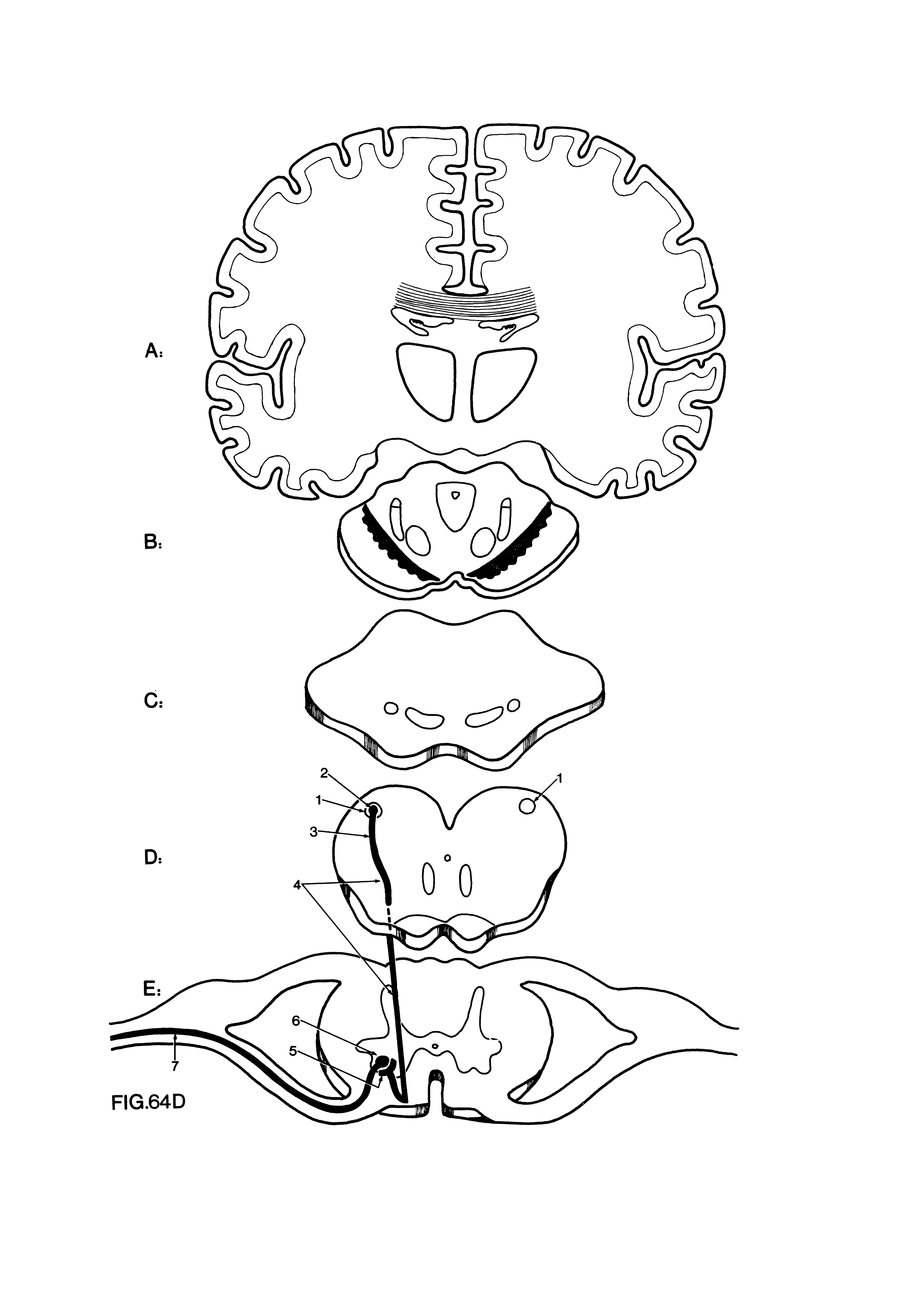
| På fosterets rygside dannes en længdegående fure, neuralfuren (1) i det yderste kimblad, ektoderm (K). | |
| Den er i begyndelsen flad, men dybden tiltager samtidig med, at randene vokser mod hinanden. Fra randene afsnøres på hver side af midtlinien en cellestreng, crista neuralis (2). | |
| Herfra udvikles bla sensitive og autonome nervecelle ganglier (“knuder”). Når neuralfurens rande vokser sammen, dannes neuralrøret (N), der adskilles helt fra overfladen. | |
| Lumen i neuralrøret (N) er spalteformet, sammentrykt fra side til side. På hver sidevæg i røret dannes en længdegående fure, sulcus limitans (3). | |
| Sulcus bliver dybere og opdeler derved neuralrørets sidevæg i en alarplade, lamina alaris (4), der ligger bagtil = dorsalt og en basalplade, lamina basalis (5), der ligger fortil = ventralt. | |
| I alarpladen dannes grå substans (sort), som modtager nerveimpulser fra receptorer. Nervetrådene kaldes afferente eller sensitive. I basalpladen dannes grå substans (L & F) eller (P & M) hvorfra nerveimpulser til effektorer afgår. | |
| Disse nervetråde kaldes efferente. Nervetråden (6) går til en tværstribet muskelcelle, der er underkastet viljens konrol. Den kaldes derfor somatisk efferent. Nervetråden (7) løber til en kirtel eller glat muskelcelle og hører derfor til det autonome eller viscerale system. Den kaldes derfor visceral efferent. | |
| I begyndelsen er neuralrørets væg tynd. Den bliver tykkere efterhånden, som der dannes flere og flere nerveceller og nervetråde. Også lumen ændrer udseende. I rygmarven bliver lumen ganske lille og danner en kanal (C) midt i rygmarven. Samtidig er sulcus limitans forsvundet, og “væggen” har skiftet udseende. Den er blevet tyk og områderne med grå substans, alarplade og basalplade har fået udseende som en sommerfugl på tværsnit. Alarpladen er blevet til baghorn (sort), mens basalpladen på dette tværsnit både danner lateralhorn (L) og forhorn (F). |
Side 297
| I hjernestammen, fx den forlængede marv, bliver væggen også tykkere, og sulcus limitans (3) bevares. “Loftet” (tyk pil) forbliver derimod tyndt, og lumen (V) fortsætter med at være ret stort i denne del af hjernestammen. | |
| De efferente nervetråde (6) og (7) kommer fra multipolare nerveceller i neuralrørets væg, mens de afferente (10) er fra pseudounipolare nerveceller (9) i sensitive ganglier uden for neuralrøret. Fra multipolare nerveceller i forreste del af basalpladen (F) vokser neuriterne (6) til tværstribet muskulatur (TM). Det er motoriske nervetråde. | |
| Fra multipolare nerveceller i bageste del af basalpladen (L) vokser neuriterne (7) til autonome ganglier (8), der består af multipolare nerveceller. Her danner de synapse, og neuriterne fra gangliecellen fortsætter til glat muskulatur (G) eller til kirtler (KI). | |
| I den del af neuralrøret, der bliver til hjerne og dermed også hjernestamme, splittes den grå substans op i øer, “kerner” eller “klumper” (cirkler og runde klatter). | |
| Fra en pseudounipolar nervecelle (9) i et sensitivt ganglie uden for neuralrøret vokser dendriten perifert, fx til huden (H). Fra huden fører dendriten impulser mod cellelegemet (9), og neuriten (10) fører impulser bort fra cellelegemet og centralt til grå substans udviklet fra alarplade eller basalplade. | |
| I den kranielle ende af neuralrøret dannes tre udvidelser, primære hjerneblærer (tegning til venstre). De er markeret med romertal og kaldes: | |
| I: prosencephalon, II: mesencephalon og III rhombencephalon. |
|
| Fra de primære hjerneblærer dannes fem sekundære hjerneblærer. De markeres med arabertal på tegningerne i midten og til højre. Betegnelsen prosencephalon bruges stadig om 1. og 2. sekundære hjerneblære, og rhombencephalon bruges om 4. og 5. sekundære hjerneblære. Betegnelsen mesencephalon anvendes om 3. sekundære hjerneblære, der dannes af II primære. | |
|
I: prosencephalon, II: mesencephalon og III rhombencephalon. |
|
|
1 telencephalon, storhjerne 2 diencephalon, mellemhjerne 3 mesencephalon, midthjerne 4 metencephalon, hjernebro og lillehjerne (pons og cerebellum) 5 myelencephalon, forlængede marv (medulla oblongata). |
Side 298
| Hulrummet i neuralrøret bliver for hjernens vedkommende til fire ventrikler, hjernekamre (1), (2), (4) og (5), og en kanal (3) mellem to af ventriklerne. | |
| I rygmarven bliver hulrummet til en snæver kanal, canalis centralis (10). | |
| Hjernens inddeling forstås lettest, når de enkelte hjernedeles beliggenhed angives i forhold til ventrikelsystemet. Telencephalon, storhjernen, indeholder lateralventriklerne (1). Diencephalon, mellemhjernen, indeholder tredie ventrikel (2). Mesencephalon, midthjernen, indeholder kanalen (3) mellem tredie og fjerde ventrikel. Den hedder aqueductus mesencephali, hjernens vandledning. Metencephalon og myelencephalon indeholder fjerde ventrikel (4) og (5). | |
| Under udviklingen bøjes hjerneanlægget (tegning til højre), samtidig med at telencephalon, storhjernen, vokser så meget, at den næsten dækker resten. |
| Rygmarven, medulla spinalis, er lednings- og centerorgan. Her findes sensitive og motoriske ledningsbaner samt centre, bl a for vandladning og afføring. Centerfunktionen kan bl a undersøges hos spinaldyr, hvor rygmarven skæres over lige under foramen magnum. | |
Side 299
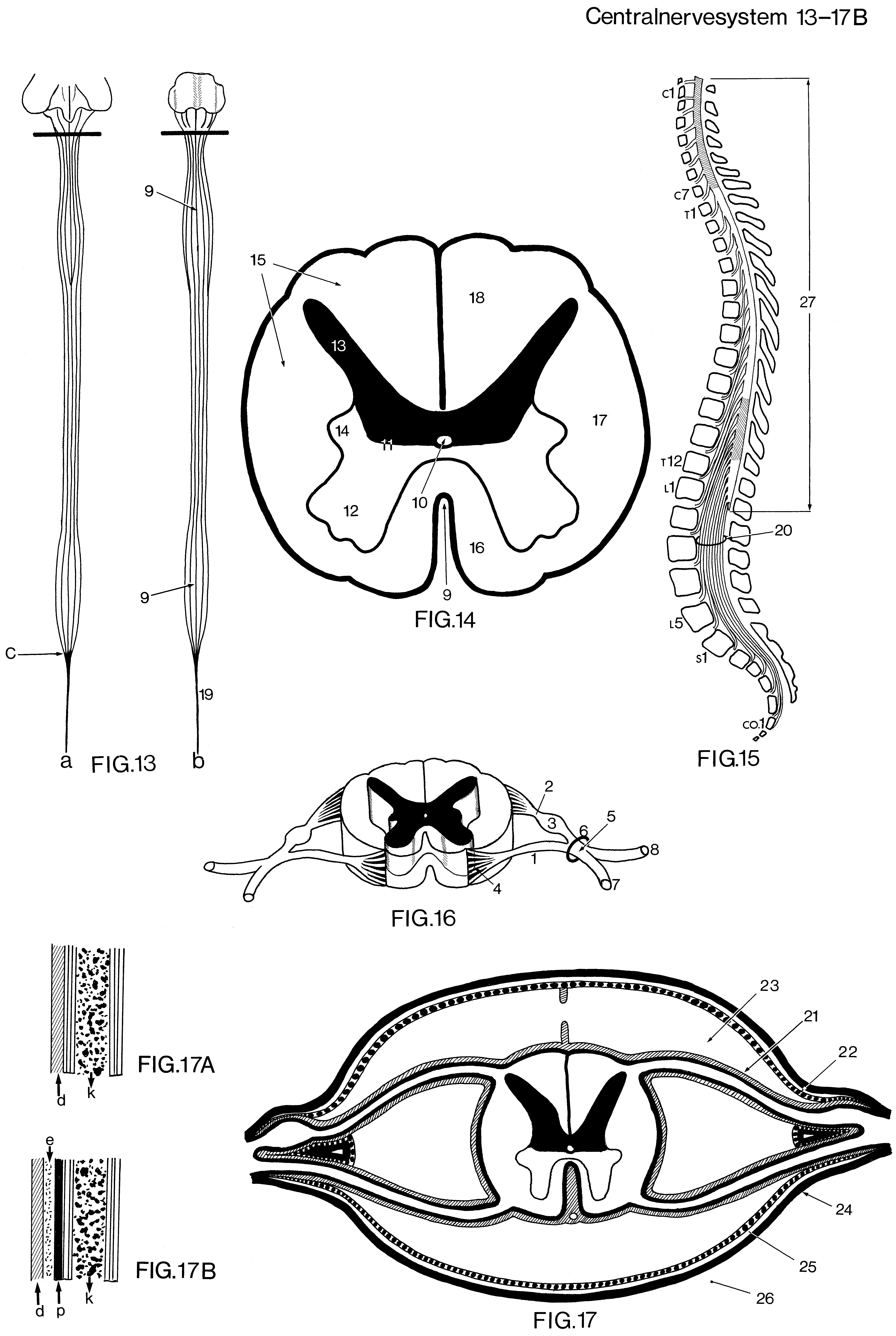
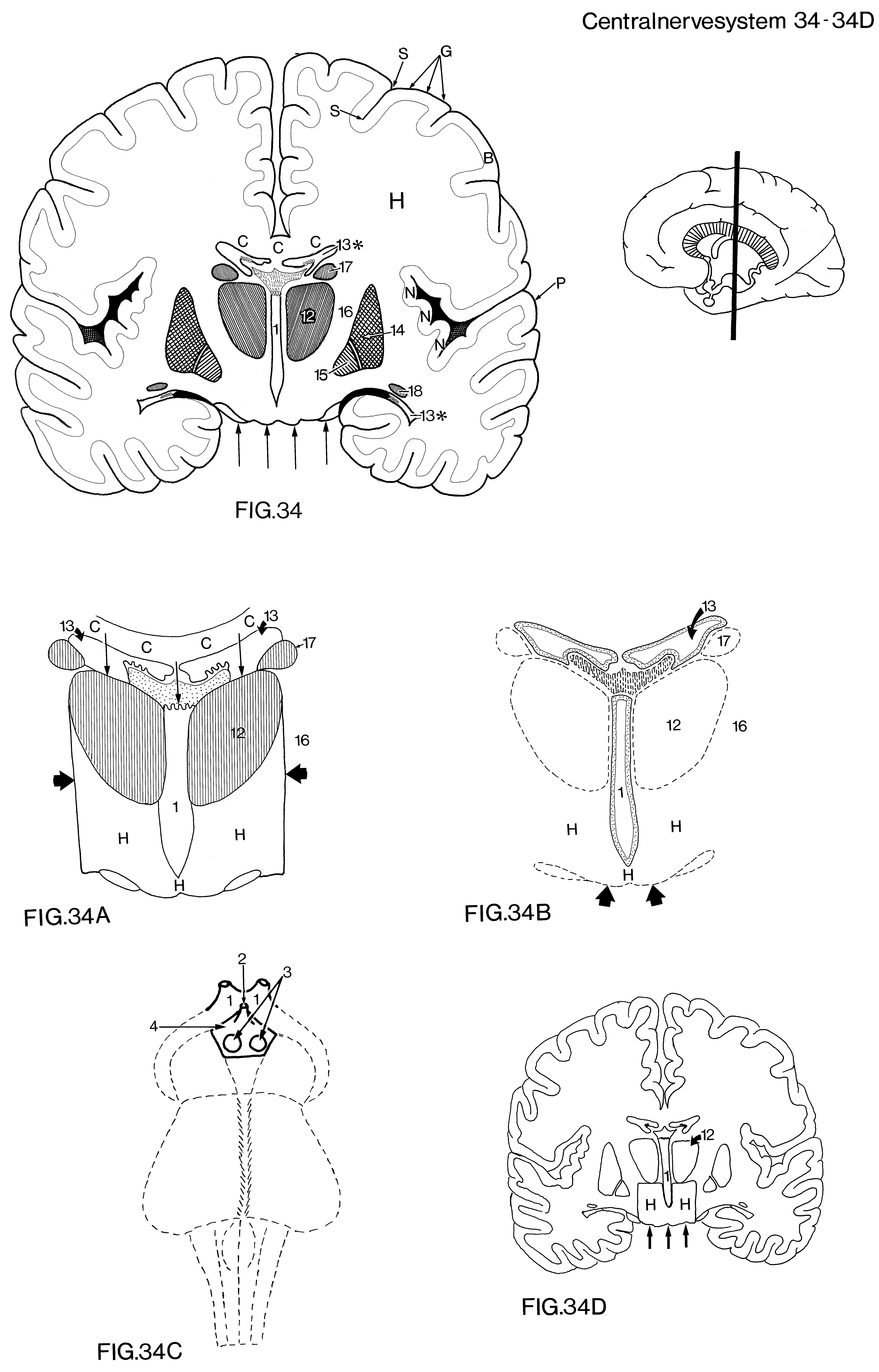
| Rygmarven (27) er en cylindrisk streng, knap en halv meter lang og en centimeter tyk, som en lillefinger. | |
| Den ender nedadtil med en kegleformet spids (C) i højde med overkanten af 2. lumbalhvirvel. Herfra fortsætter de to inderste hjernehinder som en tynd “endetråd”, filum terminale (19), til halebenet. | |
| Rygmarven er hvid og blød, og dens overflade præges af 31 par rygmarvsnerver, nn. spinales. Nerverne begynder med to rødder, radix anterior (1) fører efferente tråde, og radix posterior (2) fører afferente tråde. Udvidelsen på radix posterior er ggl. spinale (3). Begge rødder afgår med flere rodtråde, fila radicularia (4), fra rygmarvens overflade. Efter et kort forløb mødes de to rødder og danner n. spinalis (5). | |
| N. spinalis (5) forlader canalis vertebralis gennem foramen intervertebrale (6). Efter endnu et kort forløb deler nerven sig i ramus anterior (7) og ramus posterior (8). | |
| De efferente tråde (6) og (7) i radix anterior har alle trofisk centrum i multipolare nerveceller i den grå substans i medulla spinalis. De motoriske tråde (6) har trofisk centrum i forhorn og de autonome (7) i lateralhorn. | |
| De afferente tråde i radix posterior har alle trofisk centrum i pseudounipolare nerveceller (9) i spinalganglierne. Mest korrekt er at sige: Trofisk centrum er de pseudounipolare nervecellers cellelegemer, og multipolare nervecellers cellelegemer. | |
| Rygmarven består af 31 segmenter. De inddeles efter samme princip som columna vertebralis (pars cervicalis, pars thoracica osv). Ved et segment forstås en skive rygmarv med ét par rygmarvsnerver (5), en højre og en venstre, med tilhørende radix anterior (1) og radix posterior (2). Segmenterne er ikke skilt fra hinanden af tværfurer. | |
| Under udviklingen bliver medulla spinalis meget kortere end columna vertebralis. Tidligt i fosterlivet er anlæggene til medulla spinailis |
Side 300
| og columna vertebralis lige lange. Hos den nyfødte strækker medulla spinalis sig ned til 3. lumbalhvirvel. Hos en voksen ender medulla spinalis ved overkanten af 2. lumbalhvirvel. Forskydningen medfører, at rygmarvssegmenter og hvirvler kun ligger ud for hinanden opadtil. | |
| På overfladen af medulla spinalis findes seks længdefurer. | |
| Den forreste, fissura mediana anterior (9), er dybest. Den strækker sig næsten en tredjedel ind i medulla spinalis. De andre furer ses kun som svage indhak på overfladen af tværsnittet. På dette tværsnit findes iøvrigt otte furer. Det “ekstra par” bagtil findes kun i øverste del af medulla spinalis. | |
| På et tværsnit af medulla spinalis ligger canalis centralis (10) omtrent i midten. Den er omgivet af grå substans, substantia grisea, der er H-formet. H-et’s tværstreg ligger omkring canalis centralis, og på hver side består den grå substans af forhorn, cornu anterius (12) og baghorn, cornu posterius (13). I thoracaldelen findes tillige lateralhorn, cornu laterale (14). Udtrykket “horn” er udmærket, når det drejer sig om tværsnit gennem medulla spinalis.I virkeligheden er “hornene” sammenhængende søjler af grå substans. | |
| Den hvide substans, substantia alba, omgiver den grå substans på alle sider. Den hvide substans består af ledningsbaner, der er ordnet i strenge, funiculi mellem “hornene”. Strengene består af forstreng (16), sidestreng (17) og bagstreng (18). | |
| Medulla spinalis (27) ligger i canalis vertebralis. Den er fikseret ved hjælp af nn. spinales og “endetråden”, filum terminale (19), der især dannes af de omgivende hinder. | |
| Rygmarven ender ved anden vertebra lumbalis, mens hinder og de væskefyldte spalter mellem hinderne samt en del af nerverødderne fortsætter. De stærkt nedadrettede nerverødder danner en “hestehale”, cauda equina (20). | |
| Rygmarven omgives af tre hinder. Den inderste hinde, pia mater (21), beklæder rygmarvens overflade. Den mellemste hinde, spindelvævshinden, arachnoidea (22), er adskilt fra pia mater af en ret bred spalte (23), spatium subarachnoideum, der er fyldt med væske, liquor cerebrospinalis. |
Side 301
| Den yderste hinde, dura mater spinalis (24), er adskilt fra arachnoidea af en ganske fin spalte (25), spatium subdurale. I modsætning til de to blade i dura mater cranialis består dura mater spinalis kun af et enkelt “blad” eller lag. Dura mater spinalis adskilles fra væggene i hvirvelkanalen af en bred spalte, spatium epidurale (26), der er fyldt med veneplexus og fedt. Det danner et godt polster for medulla spinalis.S | |
| En vigtig relation for medulla spinalis fortil er hvirvellegemerne med bruskskiverne, disci intervertebrales. Bagtil ligger hvirvelbuerne. | |
| Hjernen er omgivet af de samme hinder og væskefyldte spalter som rygmarven, men spatium epidurale mangler i kraniehulen. Her ligger dura mater cranialis direkte på kranieknoglerne, men består til gengæld af to lag, hvor laget nærmest knoglen fortsætter i periost på udsiden, mens “det inderste” lag fortsætter som dura mater spinalis i canalis vertebralis. |
| Hjernestammen består af tre afsnit, forlængede marv, medulla oblongata (1), hjernebro, pons (2) og midthjerne, mesencephalon (3). | |
| Sammen med cerebellum danner medulla oblongata og pons rhombencephalon, der omgiver fjerde ventrikel (4) og (5). Mesencephalon er et selvstændigt hjerneafsnit omkring aqueductus mesencephali (3). Hjernestammen fungerer som ledningsorgan og centerorgan. Den indeholder sensitive og motoriske ledningsbaner og en række centre, hvoraf flere er livsvigtige. Det er centrene for åndedræt, hjertefunktion og blodtryk. Derfor er det ikke mærkeligt, at selv små læsioner af hjernestammen kan få meget alvorlige følger. | |
| Med lidt god vilje er hjernestammen nærmest cylinderformet, men overfladen er dog ret uregelmæssig med fremspring og fordybninger. |
Side 302
| Længden er ca 8 cm for hele hjernestammen, heraf udgør medulla oblongata 3 cm, pons 3 cm og mesencephalon 2 cm. Diameteren nedadtil er ca 1 cm. som medulla spinalis. | |
| Bagfladen af hjernestammen ses bedst, når cerebellum er fjernet ved at gennemskære forbindelserne (9, 10 & 11) med hjernestammen i begge sider. Herved kommer fjerde ventrikel (4) til syne. Området er rhombeformet, og det er bunden, fossa rhomboidea, der ses. Bunden skråner i retningen opad og fremad på grund af hjernestammens skrå retning, der bestemmes af clivus i fossa cranii posterior. Det er clivus, forfladen af hjernestammen vender mod. | |
| Hjernestammen er hvid på overfladen, undtagen den del der indgår i begrænsningen af fjerde ventrikel (4). Her er overfladen dækket af et lag grå substans. Området (4) hedder fossa rhomboidea. Det dannes af bagfladen af pons og øverste del af bagfladen af medulla oblongata. Hjernestammen er blød, men fastere end hjernens hemispherer. Fossa rhomboidea fortsætter opadtil i aqueductus mesencephali, der ligger i mesencephalon. | |
| Nedadtil er der forbindelse med canalis centralis (pil) i den nederste del af medulla oblongata og videre herfra i canalis centralis i medulla spinalis.Man kan også sige, at canalis centralis ender med at åbne sig i fjerde ventrikel ved nederste spids af fossa rhomboidea. | |
| Hjernestammen er beklædt med pia mater på overfladen, undtagen i fossa rhomboidea (4). Her er den grå substans dækket af et lag ependymceller mod lumen i fjerde ventrikel. | |
| På forfladen af hele hjernestammen findes en indsænkning langs midtlinien. På medulla oblongata er den smal og dyb – ligesom på medulla spinalis – og hedder fissura mediana anterior (5). | |
| På hver side af den midtstillede indsænkning findes gennem hele hjernestammen en prominens, en forhøjning, med vore vigtigste motoriske baner, pyramidebanerne. |
Side 303
| De indeholder ca 1 million aksoner, der går fra hjernebarken til medulla spinalis og er særlig vigtig for igangsætning af viljestyret bevægelse. På medulla oblongata er prominensen smal og kaldes pyramis (6). På pons er den bred og afrundet (oooo). På mesencephalon er den bred og kraftig. Her hedder den crus cerebri (flertal: crura) (7). Den overfladiske beliggenhed af vore vigtigste motoriske baner gør dem sårbare ved læsioner af hjernestammen. | |
| Hjernestammens overflade præges af udspringet af ti par hjernenerver (III-XII). Næsten alle udspringer fra forfladen, undtagelsen er hjernenerve nr. 4 (IV), der udspringer fra bagfladen af hjernestammen. Udspringene kaldes de apparente udspring. Hjernenerve nr. 3 og 4 (III) og (IV) udspringer fra mesencephalon, nr. 5 (V) fra pons, nr. 6, 7 og 8 (VI, VII, VIII) udspringer på overgangen mellem pons og medulla oblongata, og de fire sidste hjernenerver 9, 10, 11 og 12 (IX, X, XI, XII) udspringer fra medulla oblongata. | |
| Hjernenerve nr. 4 (IV) udspringer på bagfladen af mesencephalon lige under (8). | |
| Ved en nerves apparente udspring forstås det sted på centralnervesystemet, hvor den kommer til syne. Her forlader den hjernens eller rygmarvens substans og bliver fri. Hjernenerve nr. 1 og 2 udspringer ikke fra hjernestammen, og de er også på andre måder ret specielle. Modsat spinalnerverne har hjernenerverne ingen klar deling mellem motoriske og sensoriske rødder. Trådkomponenterne er oftest blandet sammen allerede ved nervens apparente udspring. | |
| Hjernestammens tre dele (1), (2) og (3) kan let afgrænses fra hinanden på forfladen. Overgangen mellem medulla oblongata og medulla spinalis er jævn. Grænsen sættes ved nedre kant af foramen magnum. Hjernestammens øverste del fortsætter uden skarp overgang i diencephalon ved de to cirkler mellem crus cerebri. Cirklerne viser corpora mamillaria, der hører til diencephalon. | |
| På bagfladen af hjernestammen er grænsen mellem pons og medulla oblongata utydelig. Det skyldes, at begge indgår i fossa rhomboidea (4), og her er ingen tværfure til at vise grænsen. På overfladen af medulla oblongata, under fossa rhomboidea, findes flere prominenser (pile) adskilt af længdegående furer. | |
| Prominenserne dannes af underliggende bagstrengskerner, dvs øer af grå substans med forbindelse til banerne i bagstrengen i medulla spinalis. På bagfladen af mesencephalon ligger fire afrundede fremspring, firhøjene (8). Der er to øvre og to nedre firhøje. | |
| Firhøjene er karakteristiske for bagfladen af mesencephalon. | |
| Hjernenerve nr. 4 (IV) udspringer på bagfladen af mesencephalon lige under (8). |
Side 304
| Hjernestammen og cerebellum er forbundet ved hjælp af tre lillehjernestilke, pedunculi cerebellares, i hver side. Stilkene (9), (10), (11) er skåret over for at fjerne cerebellum. Cerebellum står i forbindelse med mesencephalon gennem pedunculus cerebellaris superior (9), med pons gennem pedunculus cerebellaris medius (10), der er den tykkeste, og med medulla oblongata gennem pedunculus cerebellaris inferior (11). Hjernestammen kan kun skilles fra cerebellum ved at skære de tre stilke over. | |
| Et tværsnit gennem nedre del af medulla oblongata minder om et snit gennem medulla spinalis. I denne del af medulla oblongata findes canalis centralis (pil) stadig. Men den grå substans har ændret form. | |
| På et tværsnit gennem øvre del af medulla oblongata er der sket endnu flere ændringer. Canalis centralis mangler og er erstattet af fossa rhomboidea (pil). Den grå substans er samtidig blevet splittet op i mindre områder, som vi kalder kerner (sorte, hvide, skraverede). Kernerne varierer meget i størrelse. | |
| Et tværsnit gennem pons ligner et snit gennem øvre del af medulla oblongata. | |
| På et tværsnit gennem mesencephalon ses aqueductus mesencephali (pile) omgivet af grå substans (lodret skravering). Endvidere findes særlige kerneområder, “den sorte substans”, substantia nigra (12), og “den røde kerne”, nucleus ruber (13). Den del af mesencephalon, der ligger dorsalt for aqueductus mesencephali, er tagpladen (14), med firhøjene (8). | |
| Den del af mesencephalon, der ligger ventralt for aqueductus mesencephali, er “storhjernestilken”, pedunculus cerebri (15). Mellem |
Side 305
| højre og venstre crus cerebri findes en fordybning, fossa interpeduncularis, med det apparente udspring af hjernenerve nr. 3. | |
| Både nucleus ruber og substantia nigra består af nerveceller, og de hører dermed til hjernestammens “grå” substans trods deres røde og sorte farve. Den grå substans (lodret skravering) omkring aqueductus mesencephali er forsynet med et par små hvide felter nedadtil (pilehoved). Det er kernen for hjernenerve nr. 3 i højre og venstre side. |
| Hjernestammen ligger i fossa cranii posterior opad clivus (venstre pil fra 3). Hjernestammen er fikseret gennem forbindelsen med medulla spinalis nedadtil, og med diencephalon og telencephalon opadtil. | |
| Endvidere er den fikseret ved hjælp af tre stilke til lillehjernen (9), (10), (11). | |
| Endelig holdes hjernestammen fast af de nerver (III-XII), der udspringer fra den, og som går gennem dura mater i basis cranii. | |
| Den vigtigste relation for pons og øvre del af medulla oblongata (4) er fjerde ventrikel (1) og cerebellum (3). Den store tegning svarer til “rammen” på den lille tegning. |
| Med en enkelt undtagelse er hjernen omgivet af de samme hinder og spalter som medulla spinalis, dvs dura mater (2), arachnoidea (4), pia mater (6), spatium subdurale (3) og spatium subarachnoideum (5). I cavitas cranii mangler spatium epidurale. Her beklæder dura mater (2) knoglernes indside (1). | |
| I kraniehulen ligger dura mater direkte på knoglernes indside og fungerer som periost. I canalis vertebralis er dura mater adskilt fra periost (fig. 17 B) på hvirvlerne af fedt og veneplekser i spatium epidurale (e). Spatium subdurale (fig. 26 B & C) (3) er en ganske smal spalte. | |
| I subarachnoidalrummet, langs hjernestammens forflade, ligger arterier. A. vertebralis sin. og dxt. (18) gennemløber halshvirvlernes tværtappe og fortsætter gennem foramen magnum ind i cavitas cranii. Her løber de sammen og danner én stor arterie, a. basilaris (19), der ligger i den brede sulcus basilaris midt på forfladen af pons. | |
| Selv om der ikke findes spatium epidurale i kraniehulen, løber dura maters arterier (5) mellem knogle (1) og dura (2). | |
Side 306
| Fjerde ventrikels skråtstillede bund er fossa rhomboidea (4). Den dannes af bagfladen af pons opadtil og af øvre del af medulla oblongata’s bagflade nedadtil. Loftet dannes af cerebellum, der her er fjernet. | |
| Fjerde ventrikel (4) har opadtil forbindelse med tredie ventrikel (7) gennem aqueductus mesencephali (6). Nedadtil er der direkte forbindelse fra fjerde ventrikel til canalis centralis (2) og åbninger til subarachnoidalrummet (pile). | |
| Set på et mediansnit er fjerde ventrikel (4) teltformet, og spidsen af teltet (stjerne) vender bagud mod cerebellum (8). Den del af cerebellum, der ligger under spidsen, dækkes af tela choroidea (5). Det består af ependym (sort), bindevæv (pia mater) (prikket) og talrige kar. Her produceres en del af cerebrospinalvæsken, liquor cerebrospinalis. | |
| Liquor cerebrospinalis forlader fjerde ventrikel gennem tre huller i tela choroidea, i nederste del af loftet. Det midterste af de tre huller ses ved spidsen af to pile. Hullerne fører til subarachnoidalrummet, spatium subarachnoideum, hvor stjernerne står. | |
Side 307
| Overskydende cerebrospinalvæske tømmes over i hjernens blodsinus (10) gennem udposninger (9). Udposningerne består af arachnoidea og dura mater. |
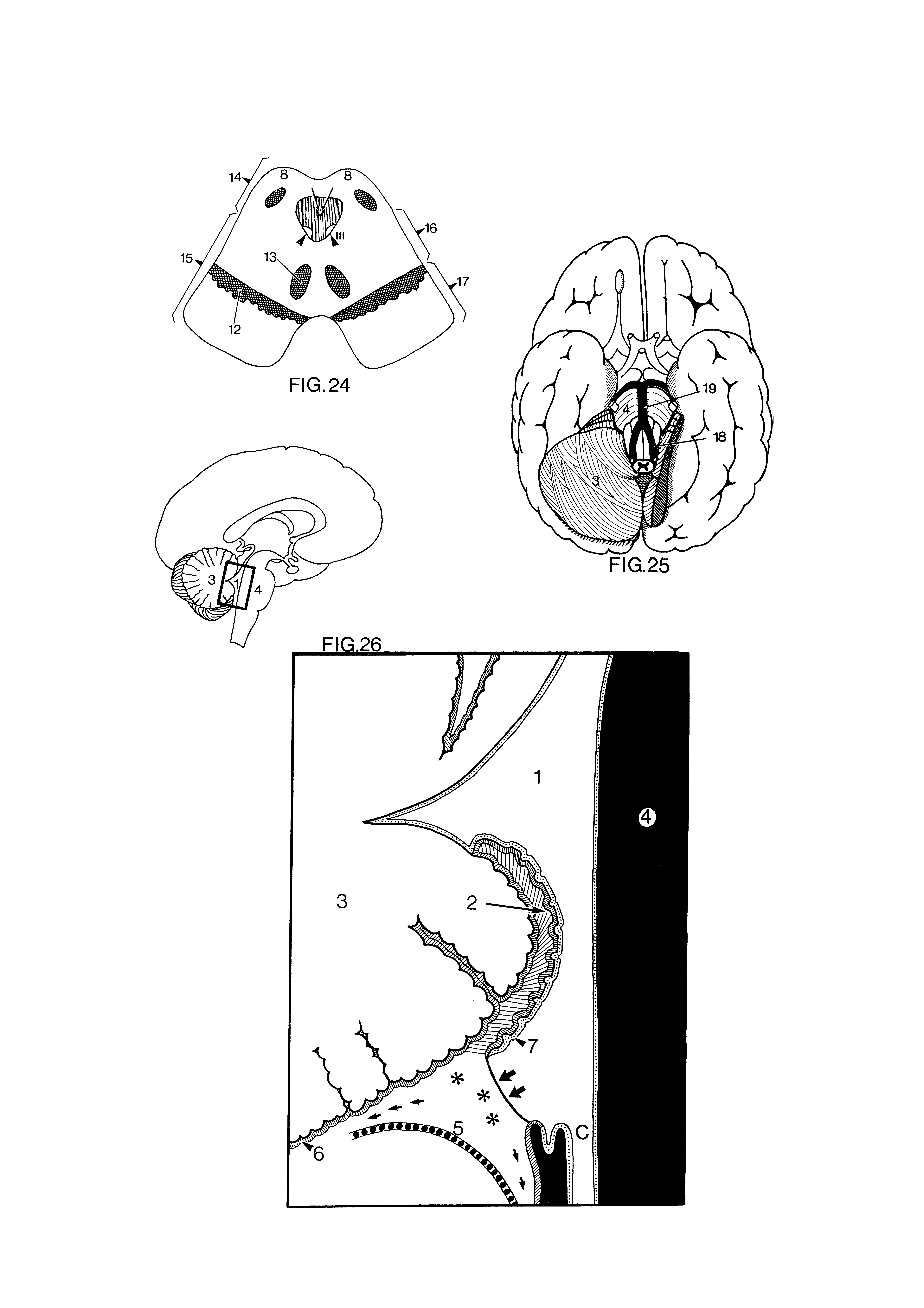
| I fossa rhomboidea er sulcus limitans (3) bevaret mellem alarplade og basalplade. I alarpladen i fossa rhomboidea dannes hjernenervekerner (sorte), der svarer til baghorn i rygmarven, og i basalpladen i fossa rhomboidea dannes kerner (skraveret og hvide), der svarer til lateralhorn og forhorn i rygmarven. | |
| I den mediale del af basalpladen findes motoriske hjernenervekerner (hvide), der svarer til forhorn. I den laterale del af basalpladen, lige medialt for sulcus limitans (3), ligger autonome (skraverede) hjernenervekerner, der svarer til lateralhorn. Lateralt for sulcus limitans, i alarpladen, ligger sensitive hjernenervekerner (sorte i venstre side). De svarer til baghorn. Kernernes beliggenhed svarer stort set til hjernenervernes apparente udspring fra hjernestammen. | |
| Hjernenervekernernes indbyrdes beliggenhed fremgår tydeligt, når de tegnes ind på en hjernestamme set bagfra. De sorte kerner er sensitive, de hvide er motoriske og sender impulser til tværstribet muskulatur, mens de skraverede er autonome og her af typen parasympatiske. De sender impulser til kirtler eller glat muskulatur. Der findes naturligvis sensitive og motoriske samt autonome kerner i begge sider, men for overskuelighedens skyld er her kun tegnet sensitive (sorte) til venstre og motoriske og autonome til højre. | |
| Kernernes tal svarer til hjernenervernes numre. Hjernenerve nr. 5 har en sensitiv kerne (sort), der er mange centimeter lang, idet den strækker sig gennem hele hjernestammen. Hjernenerve nr. 5 har tillige en motorisk kerne (til højre), men den har en mere beskeden størrelse. |
Side 308
| Hjernenervekerner ligger i forskellig dybde i hjernestammen. Det ses, når kernernes placering projiceres ind på et midtsagittalsnit. Det er fjerde ventrikel (IV) til venstre. Ligevægtsnervens kerner, vestibularkerner, (nederste 8) ligger tæt på bunden af fjerde ventrikel. Øverste (8) er hørenervens kerne(r). | |
| De motoriske (M) og autonome – der her er parasympatiske – (P) hjernenervekerner er udspringskerner (U) for motoriske og autonome, parasympatiske nervetråde. Kernerne består af multipolare nerveceller. Cellelegemerne i de motoriske kerner er trofisk centrum for motoriske tråde (6) til tværstribet muskulatur. Cellelegemerne i de autonome, parasympatiske kerner er trofisk centrum for parasympatiske tråde (7) til glat muskulatur og kirtler. | |
| Både motoriske og parasympatiske nervetråde kan løbe i den samme hjernenerve, men de opfører sig forskelligt. De motoriske tråde (6) går direkte til den tværstribede muskulatur (TM). Der er kun et enkelt neuron involveret. De autonome, parasympatiske tråde (7) går til et perifert autonomt parasympatisk ganglie, hvor de danner synapse med en ny multipolar nervecelle (8), som bliver trofisk centrum for aksonen til glat muskulatur (G) og kirtler (KI). Forløbet fra udspringskerne involverer således to neuroner. | |
| De autonome kerner i hjernestammen er parasympatiske. Lateralhornene i medulla spinalis er også autonome, men de tilhører den sympatiske del af det autonome system. Under beskrivelsen af det autonome nervesystem bliver der nærmere gjort rede for forskellen mellem de to dele. | |
| Sensitive hjernenervekerner (S) er terminalkerner (T) og består af multipolare nerveceller. Cellerne er ikke tegnet, men de danner synapse med neuriter (10) fra pseudounipolare nerveceller (9), der ligger med deres cellelegeme i sensitive hjernenerveganglier. Her er således trofisk centrum for hjernenervernes sensitive tråde. Hjernenerveganglier svarer til spinalganglier,der ligeledes indeholder pseudounipolare nervecellelegemer (9). | |
| Under beskrivelsen af mesencephalon nævntes substantia nigra og nucleus ruber, der begge er eksempler på andre kerneområder i hjernestammen. Herudover findes gennem hele hjernestammen utallige |
Side 309
| små kerner spredt mellem de langt større hjernenervekerner. De benævnes under ét formatio reticularis. Denne struktur har afgørende betydning for bevidsthed og for musklernes spændingstilstand, der kaldes tonus. Den funktionelle betydning er således meget stor og omtales senere. |
|
medulla oblongata og pons.
Rhombencephalon inddeles i metencephalon, der består af pons og cerebellum, og myelencephalon, medulla oblongata.
Cerebellum har betydning for ligevægt, muskeltonus og muskelkoordination. |
|
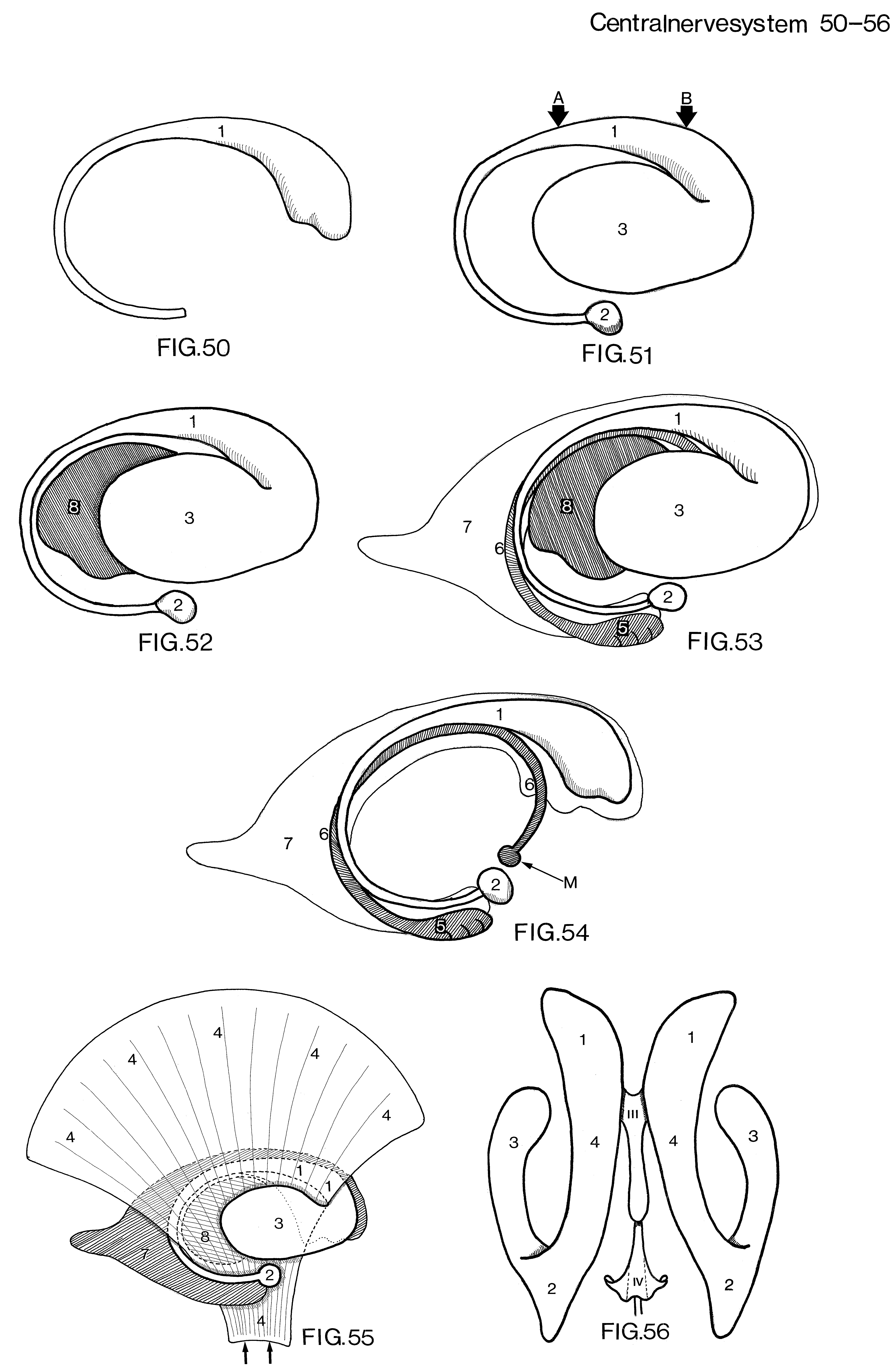
| Lillehjernen ligner lidt et stort æg med den lange akse fra side til side. Vægten er 150 gram. Farven er grå og konsistensen blød. Overfladen præges af talrige tynde lillehjernevindinger, der er adskilt af smalle dybe furer. Overfladen er beklædt med pia mater. | |
| På snitfladen ses yderst cortex cerebelli (1), lillehjernebark, der består af grå substans. Under cortex findes hvid substans (2) med nogle centrale kerner, dvs øer af grå substans. | |
| Den største af kernerne er nucleus dentatus (3). Nærmere midtlinien og loftet af fjerde ventrikel ligger en mindre kerne, nucleus fastigii (7). | |
| Cerebellum (C) ligger i fossa cranii posterior, bag pons (10) og medulla oblongata (11). Den danner loft (pile) i fjerde ventrikel (9). | |
| Lillehjernen fikseres til hjernestammen af tre pedunculi cerebellares, lillehjernestilke (5), i hver side. Lillehjernestilkene er tykke bundter af nervetråde. | |
| Pedunculus cerebellaris superior (2) forbinder lillehjerne og mesencephalon (1). | |
| Pedunculus cerebellaris medius (3) er størst og forbinder lillehjerne (4) og pons (1), mens | |
| pedunculus cerebellaris inferior (5) forbinder lillehjerne (3) med medulla oblongata (4). |
| Cerebellum ligger i fossa cranii posterior, og dens midterste afsnit (3) danner loft (eller bagvæg) i fjerde ventrikel (1). Loftet dækkes nedadtil af tela choroidea (2 og 7), men er ellers beklædt med ependym som resten af fjerde ventrikel. |
Side 310
| Ovenover cerebellum danner dura mater tentorium cerebelli (3), der skråner lidt til siderne fra midtlinien. Tentorium spænder sig mellem sulcus sinus transversi og bageste, øvre kant på pars petrosa. Det skiller derved cerebellum fra storhjernens bageste og nederste del, der her repræsenteres af lobi occipitales. |
| Cerebellum har to flader, en øvre og en nedre. Størstedelen af begge flader dannes af hemisphererne (H), en højre og en venstre, der forbindes af et smalt midterparti, vermis (4). Øvre flade af cerebellum følger underfladen af tentorium cerebelli. | |
| Den midterste del af nedre flade, der danner loft i fjerde ventrikel, består af to skråninger, en øverste (én pil) og en nederste (to pile), som mødes i en spids (pilehoved). Ud for spidsen ligger nucleus fastigii (F). | |
| De laterale afsnit (H) af nedre flade på cerebellum passer til de flade fordybninger i kraniebunden, som ligger lateralt for foramen magnum. Endelig præges nedre flade af de tre pedunculi cerebellares (5). Den vigtigste inddeling af cerebellum refererer til, at lillehjernen består af tre afsnit med forskellig funktion og forskellig “alder”, fordi de er opstået på forskellige tidspunkter i udviklingshistorien (fylogenesen). | |
| Archicerebellum (8) er det ældste og mindste afsnit. Det har betydning for ligevægten. Paleocerebellum (6) er yngre og større og har betydning for udførelse af grove bevægelser bl a gang, mens neocerebellum (7) er yngst og størst og har betydning for udførelse af fine bevægelser som for eksempel fingerbevægelser. | |
| Barken er opbygget ens i de tre lillehjerneafsnit. Den består af tre lag, af hvilke det midterste indeholder Purkinjeceller (P). Det er multipolare neuroner med flaskeformede cellelegemer. Fra den brede ende af cellelegemet afgår neuriten. Den går til en af cerebellums kerner (K). Fra archicerebellum er det til nucleus fastigii (7). Som det ses, afgår purkinjecellens dendriter fra den smalle ende af cellelegemet. De forgrener sig “espalieragtigt” i ét plan, og de bliver i barken. |
| Gennem pedunculus cerebellaris inferior (stor pil) modtager archicerebellum (3) tråde (2) fra vestibularkernerne (1) i medulla oblongata, og sender tråde (5) tilbage ad samme vej til vestibularkernerne, via nucleus fastigii (7), der er knyttet til archicerebellum. |
Side 311
| Vestibularkerner er ligevægtsnervens kerner, og den vigtigste er Deiters kerne. | |
| Den reciprokke forbindelse (2) og (5) mellem cerebellum og vestibularkerner viser, at de er tæt knyttet sammen. Impulser fra buegange og forgårdssække i det indre øre føres centralt til vestibularkernerne af n. vestibularis, ligevægtsnerven. Her kan de hurtigt omkobles til archicerebellum. | |
| Ved opretholdelse af ligevægten arbejder ligevægtsorganerne i det indre øre, vestibularkerner og archicerebellum sammen. Desuden sender vestibularkernnerne (1) impulser til medulla spinalis igennem en ledningsbane (6). Denne bane løber i forstrengen af medulla spinalis, og trådene her danner synapse med forhornsceller, som på den måde kan påvirkes af ligevægtsapparatet. Vestibularkernerne har også forbindelse med hjernenervekerner til øjenmuskler og med kerner for muskler, der bevæger hovedet (ikke vist). Gennem ligevægtsapparatet er der således mulighed for at påvirke øjets bevægelser samt stilling af hovedet og den øvrige del af kroppen. Fra archicerebellum er desuden forbindelse (4) med formatio reticularis (8) i hjernestammen. | |
| Paleocerebellum (2) modtager tråde fra medulla spinalis gennem to forskellige ledningsbaner (1b og 1a). Fra paleocerebellum (2) går der tråde via nucleus dentatus (3) til nucleus ruber (5) og thalamus (7) i diencephalon. | |
| Disse ledningsbaner forsyner lillehjernen, cerebellum, med oplysninger om bl a musklernes kontraktionstilstand, stilling af arme og ben og lignende, der under et kaldes for proprioceptive impulser eller dybdesensibilitet. | |
| Neocerebellum (4) modtager impulser fra forskellige afsnit af cortex cerebri (1). Det sker gennem ledningsbaner (2), der omkobles i pons (3), hvor der er synapse. |
Side 312
| Fra neocerebellum (4) sender Purkinjeceller impulser tilbage gennem nucleus dentatus (6) og thalamus (7) til den motoriske del af cortex cerebri. Når cortex cerebri sender impulser til tværstribet muskulatur, skeletmuskulatur, og denne starter en bevægelse, så modtager cerebellum samtidig information gennem ledningsbaner fra hjernebarken. Samtidig ved vi nu, at cerebellum hele tiden modtager en strøm af informationer, der sørger for, at cerebellum er nøje orienteret om musklernes kontraktionstilstand, legemsdelenes indbyrdes placering, hovedets stilling, trykket på hudoverfladen mv. | |
| Alle informationerne gør cerebellum i stand til at registrere bevægelser, der rent faktisk sker, og sammenligne dem med de bevægelser, der var ønsket fra cortex cerebri. Cerebellums vigtigste opgave er, at gøre forskellen mellem de ønskede og de faktiske bevægelser så lille som mulig. Det sker ved at cerebellum sender korrigerende signaler både til cortex cerebri via thalamus og til kernecentre for de ledningsbaner, der fører aktionspotentialer til motoriske celler i forhorn eller hjernenervekerner. | |
| Cerebellum har forbindelse med andre kerner i hjernestammen end de før nævnte.Talrige tråde (3) kommer fra formatio reticularis (1) og går både til archi-, paleo- og neocerebellum. | |
| Cerebellums vigtigste udfaldsport er nucleus dentatus. Den modtager tråde fra Purkinjeceller i både neo- og paleocerebellum. Trådene fra paleocerebellum (2) går til nucleus ruber (5) og thalamus (7). Fra nucleus ruber er der forbindelse til medulla spinalis gennem ledningsbaner. | |
| Alle tråde fra neocerebellum til storhjernen går via thalamus (7) til den motoriske del af cortex cerebri, area 4 og 6. | |
| For at få overblik over forbindelserne, kan cerebellum (II) betragtes som et stort reflekscentrum, der modtager og behandler impulser fra sensitive og motoriske områder, og som afsender impulser til motoriske områder.
Cerebellum (II) får impulser fra (I) cortex cerebri (1) via pons, kerner i hjernestammen vestibularkerner (2), formatio reticularis (3), og fra medulla spinalis (5) gennem ledningsbaner. |
Side 313
| Cerebellum (II) sender impulser til (III) motorisk cortex (via thalamus) (6) og til kerner i hjernestammen nucleus ruber (7), vestibularkerner (8) og formatio reticularis (9). Fra kernerne i hjernestammen ledes impulser til forhorn i medulla spinalis gennem ledningsbaner. |
| Cerebellum er vigtig for opretholdelsen af ligevægten (1). Svimmelhed (2) ses ved lidelser i cerebellum, især i archicerebellum. | |
| Cerebellum er vigtig for den normale muskelkoordination (3) og (4). Særlig nødvendigt er samarbejdet mellem musklerne ved komplicerede bevægelser, og her har lillehjernens styring afgørende betydning. Muskelkoordination kan undersøges ved “finger-til-næse-prøven”. Patienten lukker øjnene og opfordres til at føre en finger til sin egen næsetip (3). Ved lidelser i lillehjernen bevæges fingeren i en uregelmæssig bane, til den når målet (4). Patienten kan ikke standse præcist ved næsetippen, men presser og gnider fingeren mod næsen. | |
| Manglende muskelkoordination kan også afsløres ved at lade patienten udføre bevægelser, der hurtigt afløser hinanden, fx hurtig supination og pronation af underarmen. Normalt kan disse bevægelser udføres meget hurtigt, men hos patienter med en cerebellarlidelse bliver bevægelserne langsomme og usikre. Muskelkoordination kan også prøves ved at lade patienten føre to fingre mod hinanden som vist på (5). Ved svigtende muskelkoordination bliver resultatet som vist på (6). | |
| Ved cerebellarlidelser optræder endvidere tonusnedsættelse, idet en normalt fungerende lillehjerne er nødvendig, for at opretholde musklernes normale spændingstilstand, tonus. Muskeltonus undersøges ved, at lade patienten holde armene parallelt foran sig med lukkede øjne og derefter bevæge armene parallelt bort fra hinanden. Normalt kan armene holdes parallelt (7), også med lukkede øjne. Ved lidelser i cerebellum devierer armen i den syge side (8). |
Side 314
| Nedsat tonus viser sig også i patientens gang. På grund af nedsat tonus i den ene side afviger gangen hele tiden fra den lige linie. Når afvigelsen erkendes, ændres retningen straks. Resultatet bliver “fuldemandsgang” i zigzag. | |
| Nystagmus ses hos patienter med cerebellarlidelser og er et andet udtryk for tonusnedsættelse. Ved nystagmus udfører øjnene rytmiske bevægelser, der består af to faser, en langsom, hvor øjnene “glider” bort fra fikspunktet, og en hurtig, hvor de føres tilbage til udgangsstillingen, hvorfra de igen glider langsomt bort. Og således fortsætter øjenbevægelserne rytmisk. | |
| Endelig kan lidelser i cerebellum give sig udslag i intentionstremor, dvs “rysten på hånden”, når der skal udføres tilsigtede bevægelser, fx løfte et glas, mens andre former for bevægelse, der er reflektorisk betingede, udføres næsten normalt. |
| Mellemhjernen, diencephalon, omfatter tredie ventrikel (1) med omgivende strukturer. På mediansnittet her er tredie ventrikel (1) ramt i hele sin udstrækning, forfra – bagud og op – ned. Pilene peger på begrænsningen. | |
| De tykke sidevægge i diencephalon kaldes thalamus (12), og bunden betegnes hypothalamus (H’er). | |
| Under hypothalamus ligger hypofysen (7), der er kroppens overordnede endokrine kirtel. | |
| Thalamus spiller en central rolle som koblingsstation for sanseinformation til storhjernen. Bortset fra nervebanerne med lugteimpulser, løber alle sensoriske nervebaner via centre i thalamus, før de ender i cortex cerebri. | |
| Hypothalamus virker som bindeled mellem nervesystemet og det endokrine system. Denne del af hjernen kontrollerer de fleste endokrine kirtler i kroppen enten direkte eller indirekte. Den direkte kontrol sker ved, at hypothalamus producerer en række forskellige hormoner, som fremmer eller hæmmer de forskellige endokrine celler i hypofysens forlap. Desuden producerer hypothalamus to hormoner, som frigøres fra hypofysens baglap. |
Side 315
| Den indirekte kontrol sker ved, at hypofysen styrer aktiviteten i en række af de andre endokrine kirtler. | |
| Hypothalamus virker også som et overordnet kontrollerende center for det autonome nervesystem og har en central rolle i kontrollen af legemstemperaturen. Desuden er følelser som tørst, sult og seksualdrift knyttet til bestemte områder i hypothalamus. Det samme er følelsen af velbefindende, når disse drifter er tilfredsstillet. | |
| Sammen med hjernestammen er hypothalamus– den del af hjernen, som betyder mest for reguleringen af kroppens indre miljø. Kort sagt, diencephalon indeholder overordnede autonome centre, er mellemstation for store afferente baner på vej til cortex cerebri og producerer hormoner. | |
| Diencephalon er dækket af storhjerne og i stor udstrækning vokset sammen med denne. For at kunne se diencephalon som en “kasse”, må den først skæres fri. | |
| Det kan gøres ved at skære langs hele omkredsen (pile) af af tredie ventrikel (1). Diencephalon ligner så en uregelmæssig kasse med tykke vægge og spalteformet lumen. Længden er 4 til 5 centimeter. | |
| Kasseformen ses også på frontalsnit, hvor man kan lægge to lodrette snit fra (17) i hver side til hjernens nedre overflade, og dernæst forbinde (17) i højre og venstre side med et tværsnit. Så er diencephalon (12 & H) isoleret som en kasse, hvor det spalteformede lumen (1) naturligvis er tredie ventrikel. | |
| Diencephalon indeholder grå og hvid substans, og er blød. Størstedelen af overfladen er vokset sammen med telencephalon (H). Mod ventrikelsystemet er overfladen dog synlig. Den synlige eller frie overflade vender mod tredje ventrikel (1) eller mod lateralventriklerne (13). Her er den frie overflade ependymbeklædt. | |
| Diencephalon udgør også en del af hjernens underside, basis cerebri (pile). Her er den synlige overflade af diencephalon beklædt med pia mater. | |
| Den del af diencephalon, der indgår i basis cerebri, ligger over sella turcica og omfatter bl a hypofysen (7). De nærmeste relationer er kraniebunden nedadtil, mesencephalon (m) med aqueductus mesencephali (2), nedadtil og bagtil, samt telencephalon (T) opadtil og til siderne. | |
Side 316
| thalamus (12), højre og venstre, hypothalamus (H) (hypo: under) (under 1 og 12), |
|
| metathalamus (meta: efter) (23) og (24), og | |
| epithalamus (epi: over) (3). | |
| Lateralvæggen i tredie ventrikel (1) dannes af thalamus (12) og af hypothalamus (H). | |
| Bunden dannes af hypothalamus (6), (9) og (8). I bagvæggen findes indmundingen af aqueductus mesencephali (2) og tilhæftningen af “koglekirtlen”, corpus pineale (3), der hører til epithalamus. | |
| Loftet dannes af tela choroidea (4) (små pile) og forvæggen af neuralrørets endeplade, lamina terminalis (stor pil). | |
| I lateralvæggen af tredie ventrikel er der i hver side en spalteformet åbning, foramen interventriculare (5). Åbningen ligger lige foran thalamus (T) og forbinder tredie ventrikel med lateralventriklen. Foramen interventriculare er en rest af den oprindelige åbning mellem første og anden sekundære hjerneblære. Gennem foramen interventriculare strømmer liquor cerebrospinalis fra lateralventriklen til tredie ventrikel. |
| Thalamus (12) er mellemstation for store sensitive ledningsbaner. Den er ægformet og har størst udstrækning forfra-bagtil (lille tegning øverst). Længden er ca 4 cm. | |
| Thalamus er grå og blød. Den er beklædt med ependym på medialfladen mod tredie ventrikel (1) og på den del af øvre flade, der vender mod lateralventriklen (13). |
Side 317
| Lateralfladen af thalamus (12) er dækket af hvid substans med ledningsbaner (16) til og fra telencephalon. Nedre flade støder op til hypothalamus (H). | |
| Den bageste ende af thalamus er fri og beklædt med pia mater (pil, øverste lille tegning). | |
| Den grå substans udgør størstedelen af thalamus. Den danner mange kerner, der er ordnet i kernegrupper. Vi skelner mellem fire hovedgrupper, mediale (1), forreste (2), laterale (3) og bageste (4). Til den bageste gruppe regnes corpus geniculatum laterale (L) og mediale (M), dvs metathalamus. Kernegrupperne er adskilt af tynde plader af hvid substans (5). |
|
| Inden for de fire kernegrupper findes flere kernetyper. Vi nøjes med at skelne mellem associationskerner (1) og relækerner (2). | |
| Associationskerner udgør den “nyere” del af thalamus og modtager kun nervetråde/informationer fra andre thalamuskerner (dobbeltpile). De står i forbindelse (2) med associationszonerne i cortex cerebri (øverste tegning), hvor bl a sanseindtryk bearbejdes. Netop disse zoner af cortex cerebri er nye i udviklingsbiologisk sammenhæng, og de udgør et stort område af den menneskelige hjerne. | |
| Relækerner udgør den “ældre” del af thalamus. De modtager nervetråde fra kerner i centralnervesystemet uden for thalamus (1‑5) og står i forbindelse cortex cerebri (øverste tegning). Både med sensitive områder (1‑4) og den motoriske zone (5). Relækerner modtager impulser, der har at gøre med hørelse (1), syn (2), smag (3). Derudover forskellige sensitive indtryk, fx smerte- og temperaturindtryk samt dybdesensibilitet, der løber gennem rygmarv og hjernestamme (4). Endelig er der informationer fra cerebellum (5). I relækernerne omkobles nervetrådene (1‑5), dvs nervetrådene danner synapse, og impulserne fortsætter i nye neuroner til cortex. Forbindelserne mellem relækernerne i thalamus og de sensitive og motoriske områder af cortex cerebri går begge veje (6), dvs, forbindelserne mellem thalamus og cortex er reciprokke. På øverste tegning ses, at hørecentret (1) ligger i tindingelappen, synscentret (2) i nakkelappen, den sensitive zone (3‑4) i isselappen og den motoriske (5) i pandelappen. |
Side 318
| Thalamus har derudover vigtige forbindelser med flere afsnit af det særlige motoriske system, vi kalder for det ekstrapyramidale system, der bl a omfatter nucleus ruber (1), basalganglier (2) og formatio reticularis (3). | |
| Thalamus (2) har også forbindelse med hypothalamus (3). | |
| Thalamus er den store mellemstation for de sensitive ledningsbaner. | |
| Endelig betyder forbindelsen med cerebellum (5), at thalamus også er vigtig inden for det motoriske system, idet lillehjernens impulser til den motoriske del af hjernebarken passerer gennem relækerner i thalamus. | |
| Thalamus spiller en rolle for forskellige psykiske funktioner, bl a for smerteopfattelsen, som thalamus er med til at præge eller “farve”. Ved læsion af højre thalamus fx efter blødning, er det vanskeligere, at fremkalde smerte i kroppens venstre side. Når smertetærsklen først nås, er smerterne til gengæld kraftige og yderst ubehagelige. Selv et nålestik opfattes som en stærk, brændende smerte. Hos disse patienter kan der også optræde spontane smerter, som er meget stærke og temmelig upåvirkelige af smertestillende midler. |
| Hypothalamus er et lille område på hjernens underside lige under thalamus. Hypo betyder under. Hypothalamus virker som overordnet center for både det autonome nervesystem og for det endokrine system. Styringen af store dele af det endokrine system sker ved hjælp af hypofysen, som er en lille kirtel på størrelse med en ært. Den ligger under hypothalamus og er hæftet til den ved hjælp af hypofysestilken. I hypothalamus findes store autonome centre, og der foregår også hormonproduktion. Området ligner en uregelmæssig plade og vejer kun nogle få gram. |
Side 319
| Den del af øvre flade, der vender mod tredie ventrikel (1), er beklædt med ependym. Her danner hypothalamus (H) bund og en del af lateralvæggen. Resten af øvre flade dækkes af thalamus (12). | |
| Nedre flade hører til basis cerebri og er beklædt med pia mater (fire pile). Nedre flade af hypothalamus, og dermed af diencephalon, omfatter flere vigtige strukturer. | |
| Her findes synsnervekrydsningen, chiasma opticum (1), hypofysestilken (2) og to halvkugleformede forhøjninger, corpora mamillaria (3), samt tuber cinereum (4), en forhøjning mellem hypofysestilken og corpora mamillaria. | |
| Hypothalamus består af en blanding af grå og hvid substans. Den ligger over sella turcica, hvor hypofysen (7) ligger i en durasæk i fossa hypofysialis. I chiasma opticum (6) krydser de mediale tråde fra højre og venstre n. opticus. Efter krydsningen fortsætter de mediale tråde sammen med de ukrydsede laterale tråde i tractus opticus, der går bagud til metathalamus. | |
| Hypofysen (7) er et overordnet hormonproducerende organ. Den har forbindelse med resten af hypothalamus gennem hypofysestilken (9). Den ærtstore hypofyse består af to dele, en forlap der kaldes adenohypofysen (1) og en baglap, neurohypofysen (2). Forreste del af hypofysestilken (tre små pile) hører til (1) og bageste del til (2). Forlappen udvikles i fostertilstanden fra en udposning i loftet af den primitive mundhule, mens baglappen dannes af nervevæv fra området, hvor diencephalon udvikles. Efterhånden hæfter forlap sig til baglap, og sammen danner de to anlæg hypofysen. Forskellen i oprindelse forklarer, hvorfor forlap og baglap er selvstændige og uafhængige enheder. | |
| Neurohypofysen (2) består af nerveceller med glia og blodkar. Der findes ingen hormonproducerende celler. Baglapshormoner produceres af nerveceller i kerneområder (5) i hypothalamus. Herfra pakkes hormonerne i vesikler og transporteres i axonerne (B) til neurohypofysen, hvor de oplagres i axonenderne, indtil de frigøres. Efter frigørelsen diffunderer baglapshormonerne ind i baglappens kapillærer og transporteres derefter rundt i organismen med blodet. Adenohypofysen modtager ikke direkte nerveforsyning fra hypothalamus. I forlappen er der endokrine celler, rigeligt med blodkar og få nervefibre. I hypofysestilken hørende til forlappen, findes et specielt |
Side 320
| portåresystem (P), der er organiseret efter samme princip som leverens portåresystem, dvs med to kapillærnet (7 & S) koblet i serie. | |
| pothalamus. Det sker ved, at nervecellelegemerne i (8) danner regulationshormoner, der via aksonerne (A) føres til første kapillærnet (7) i hypofysestilken. Det er dannet af bl a hypofysearterien (6). Fra første kapillærnet føres regulationshormonerne gennem hypofysens portåre (P) til andet kapillærnet (S) omkring de hormonproducerende celler i forlappen. Her stimulerer eller hæmmer regulationshormonerne dannelsen og frigørelsen af forlapshormoner. Det er altså cellerne i adenohypofysen, der danner forlapshormonerne, men styringen af produktionen sker i hypothalamus. Portåresystemet er hurtigt og effektivt. Det fragter regulationshormonerne direkte fra nervecellerne i hypothalamus til forlappens endokrine celler. Derved undgår hormonerne at skulle bruge det store kredsløb for at nå frem. Det ville både medføre en tidsmæssig forsinkelse og en fortynding af hormonerne i det langt større blodvolumen. | |
| Hypothalamus har forbindelse med andre dele af centralnervesystemet. En vigtig forbindelse (4) løber i fornix (2). Den fører impulser fra hippocampus (1) i det limbiske system til corpus mamillare (3) i hypothalamus. | |
| Ved overskæring af forbindelsen eller ved læsion af hippocampus eller corpus mamillare, kan patienten ikke længere indprente sig nye indtryk og fx ikke huske, hvad man lige har læst. Derimod er man i stand til at huske begivenheder, der indtraf før læsionen. | |
| Hypothalamus (3) har desuden forbindelse med hjernebarken (1), det ekstrapyramidale system (5) og (6), thalamus (2) og som nævnt hypofysen (4). | |
| Hypothalamus er med til at regulere søvn (1), appetit (2), stofskifte (3), vandskifte (4), varme (5) og vasomotoriske reaktioner (6). | |
Side 321
| Metathalamus omfatter to små forhøjninger (23) og (24) i hver side. De ligger under den bageste frie ende af thalamus (12). Forhøjningerne hedder corpus geniculatum laterale (23) og corpus geniculatum mediale (24). | |
| Gennem nervetrådsbundter, der er synlige på overfladen, har corpora geniculata (23) og (24) forbindelse med firhøjene, de øvre (Ø) og nedre (N), på bagfladen af mesencephalon. Corpus geniculatum laterale (23) modtager tractus opticus (25) og er forbundet med øvre firhøje (Ø) i samme side. | |
| Corpus geniculatum mediale (24) er forbundet med nedre firhøje (N) i samme side. Den er synapsestation for hørebaner. | |
| Epithalamus omfatter epifysen, corpus pineale (3), og den nærmest tilgrænsende del af diencephalon. Corpus pineale (3) sidder i bagvæggen af tredie ventrikel (1) lige i midtlinien, og den er uparret. Corpus pineale indeholder kirtelceller, der producerer et hormon, der virker hæmmende på adenohypofysens produktion af kønskirtelstimulerende hormon. Ved lidelser i corpus pineale indtræder tidlig kønsmodning. |
| I forhold til andre hvirveldyr har arten menneske en række specielle egenskaber. Vi har evnen til intellektuel og kunstnerisk aktivitet, og vi har et veludviklet, abstrakt sprog. Vi har desuden en direkte, viljestyret muskelkontrol over hænderne, som er effektive griberedskaber. Menneskets egenskaber har gjort det muligt at udvikle redskaber og teknologi, komplicerede sociale netværk og det, vi kalder kultur. | |
| Disse specielle menneskelige egenskaber er knyttet til storhjernen, som hos mennesket udgør næsten 90% af hjernens vægt. Det, som i særlig grad adskiller menneskets hjerne fra hjernen hos andre pat |
Side 322
| tedyr, er størrelsen af storhjernens overflade, cortex cerebri. Hos en voksen er overfladen næsten 2400 kvadratcentimeter. Inden for kraniekassens begrænsede volumen skyldes det imponerende overfladeareal, de stærkt foldede og karakteristiske hjernevindinger. De fleste nerveceller i storhjernen ligger nær overfladen i et tykt lag af grå substans. Der findes dog også grå substans i form af centre eller kerner inden for storhjernen. De ligger blot i dybden. | |
| Cortex cerebri, hjernebarken, er det område af hjernen, som er ansvarlig for bevidste oplevelser af sanseindtryk, bevidst styring af kropsbevægelser og forskellige intellektuelle aktiviteter. Vores viden om storhjernens opgaver bygger bl a på virkningen af direkte stimulering af forskellige områder af barken. |
| Storhjernen, cerebrum eller telencephalon, består af to hemispherer. Den er nærmest ægformet med størst udstrækning forfra og bagtil og vejer ca 1200 gram. Farven er grå, og konsistensen er blød. Overfladen præges af hjernevindinger, gyri cerebri (G), og furer, sulci cerebri (S). Overfladen er overalt beklædt med pia mater (P). Yderst består telencephalon af grå substans, hjernebark, cortex cerebri (B). Under barken ligger hvid substans (H) med kerneområder af grå substans, blandt andet basale hjerneganglier (14), (15), (17) og (18). Desuden findes lateralventrikler (13). Thalamus (12) er også en ø af grå substans, men ikke et af de basale hjerneganglier. Den hører til diencephalon. | |
| Telencephalon ligger i fossa cranii anterior og media. Den bageste del hviler på durabladet, tentorium cerebelli (3), der danner “tag” over fossa cranii posterior, og dermed over lillehjernen. | |
| Telencephalon er overalt omgivet af spatium subarachnoideum (sort) med liquor cerebrospinalis. Hjernen ligger badet i liquor cerebrospinalis, der spiller en stor rolle ved at beskytte hjernen mod stød og slag. De to nederste pile viser, at liquor cerebrospinalis kommer ud i spatium subarachnoideum fra ventrikelsystemet gennem huller i loftet af fjerde ventrikel. | |
| Hjernen støttes endvidere af et sagittalt stillet halvmåneformet durablad, falx cerebri (40). | |
| Det ligger i furen fissura longitudinalis cerebri (28) og når ned til tværforbindelsen, corpus callosum (pil). | |
| Falx cerebri (2) har en konveks og en konkav kant. Den konvekse, øvre kant er hæftet til theca cranii langs sulcus sinus sagittalis superior, og her løber en vene, sinus sagittalis superior (4). Den nedre, konkave kant af falx cerebri ligger lige over tværforbindelsen mellem |
Side 323
| de to hemispherer. Her løber også en vene (5), der bagtil fortsætter i sinus rectus (6). Endelig er hjernen forankret til kraniebunden ved hjælp af hjernenerverne (III-XII), der går gennem dura mater. | |
| Telencephalon dækker og skjuler det meste af diencephalon. En del af øvre flade på thalamus (12) indgår i bunden af lateralventriklen (13) og det basale hjerneganglie, “halekernen”, nucleus caudatus (17), dækker den laterale del af øvre flade på thalamus. Lateralt for thalamus ligger capsula interna (16), et vigtigt afsnit af den hvide substans i telencephalon. | |
| elencephalons relation til basis cranii interna er vigtig. Storhjernen ligger over øjenhuler og næse- , pande- og kilebenshule. | |
| Den ligger endvidere på pars petrosa med høre- og ligevægtsorgan i det indre øre (2), (1), (3), trommehule og cellulae mastoideae. | |
| Hver hemisphere har en konveks lateralflade, en plan medialflade og en uregelmæssig nedre flade, basis cerebri. | |
| De to hemispherer er adskilt af en dyb fure, fissura longitudinalis cerebri (28), hvor falx cerebri ligger. Furen når til corpus callosum, hjernebjælken (49 skraveret), der er tværforbindelsen mellem højre og venstre hemisphere og præger hemispherernes medialflader. |
| Hver hemisphere deles i fire lapper, lobi, der er ufuldstændig afgrænset fra hinanden. Lapperne er pandelap, lobus frontalis (29), isselap, lobus parietalis (30), nakkelap, lobus occipitalis (31) og tindingelap, lobus temporalis (32). | |
| Lobus temporalis afgrænses af sulcus lateralis (33). Furen er rettet bagud og opad. | |
| Her er dog fjernet hjernesubstans fra de skraverede områder for at skaffe indblik til insula. | |
| Lobus frontalis og lobus parietalis adskilles af sulcus centralis (34). På lateralfladen af hemispheren har furen et langt forløb, og den er rettet nedad og fremad. |
Side 324
| Den (34) begynder øverst på medialfladen af hemispherene og fortsætter over deres øverste kant – omtrent midt på hjernen – til lateralfladen. Lobus occipitalis adskilles fra lobus parietalis af sulcus parietooccipitalis (35). Denne er ganske kort på lateralfladen, mens den på medialfladen løber til den fremadrettede sulcus calcarinus (36). | |
| Foran sulcus centralis ligger den motoriske vinding (zone), gyrus præcentralis (42). Bag sulcus centralis ligger den sensitive vinding (zone), gyrus postcentralis (43‑46). De sensoriske impulser fra huden til cortex cerebri ender i gyrus postcentralis, og de motoriske centre i gyrus præcentralis, som styrer skeletmusklerne, har en organisering, der svarer til de sensitive centre. De områder, der kontrollerer musklerne i ansigt og hænder, ligger fx lige overfor de centre, der modtager information fra de samme legemsdele. | |
| Hjernens overflade inddeles i “områder”, areae, der er nummererede. Gyrus præcentralis (42) er area 4 og (6, ikke vist). Gyrus postcentralis (43‑46) omfatter area 3, 1, 2 og 5. Synscentret (47) omkring sulcus calcarinus er area 17. Hørecentret (48) øverst på temporallappen er area 41. Det ligger lige under sulcus lateralis. | |
| Synscentret, area 17, i nakkelappen (2), har størst udstrækning på medialfladen af hjernen. Hørecentret, area 41, ligger i tindingelappen (3) lige under sulcus lateralis. Over sulcus lateralis, nederst på gyrus postcentralis i isselappen, ligger smagscentret (1), og på medialfladen af tindingelappen ligger lugtecentret (4). | |
| De to hemispherer er i princippet selvstændige enheder, som kontrollerer hver sin kropshalvdel. I praksis samarbejder hemisphererne dog om at løse forskellige opgaver, så individet fungerer som en enhed. Det vigtigste anatomiske grundlag for dette samarbejde er hjernebjælken, corpus callosum (49), der ses, når de to hemispherer trækkes fra hinanden. Den ligger i bunden af den midtstillede fure, sulcus longitudinalis, mellem de to hemispherer. Corpus callosum er hjernens største commissurbane. Den består af mere end 200 millioner nervetråde, der forbinder de to hemispherer. Gennem dette massive nervebundt går der ca 4 milliarder nerveimpulser i sekundet. |
Side 325
| På et mediansnit af hjernen (øverst) har snitfladen af corpus callosum (49) form som en C-formet “bjælke” med konkaviteten nedad, og dens forreste og bageste ende er bøjet. Lateralt er det umuligt at sige, hvor corpus callosum holder op. På frontalsnittet nederst ses, at dens utallige tråde blander sig med hvid substans i hemisphererne (H). | |
| Under corpus callosum ligger septum pellucidum (50). Denne plade adskiller de to lateralventrikler fra hinanden. Under corpus callosum bagtil og under septum pellucidum fortil ligger fornix (51), en vigtig ledningsbane. | |
| På et frontalsnit ses septum pellucidum (S) som en skillevæg mellem de to lateralventrikler (13). Septum pellucidum forbinder corpus callosum (C) med fornix (F). Da fornix har en kraftigere krumning end corpus callosum, vil højden på septum pellucidum variere efter, hvor langt fremme eller tilbage frontalsnittet ligger. Det er også sådan, at da septum pellucidum kun har en begrænset udstrækning i retning forfra-bagud, vil det kun danne skillevæg for den centrale del af lateralventriklerne. |
| Det er tidligere blevet nævnt, at den grå substans i storhjernens hemisfærer ligger som større eller mindre kerner eller områder omgivet af hvid substans. De største områder med grå substans er de såkaldte basalganglier, der har vigtige opgaver, når det gælder kontrol af bevægelser. De modtager mængder af forbindelser fra cortex cerebri og har især forbindelser retur til de motoriske dele af cortex cerebri. | |
| De vigtigste basale hjerneganglier eller basalganglier er “halekernen”, nucleus caudatus (17) og (18), “linsekernen”, nucleus lentiformis, der består af putamen (14) og globus pallidus (15) og | |
| “mandelkernen”, corpus amygdaloideum (2). De regnes alle med til den del af det motoriske system, vi kalder det ekstrapyramidale system. |
|
| Nucleus caudatus (1) ligner et stærkt bøjet komma. Konkaviteten i kommaet vender fremad. Den tykke del af nucleus caudatus kaldes caput og ligger i frontallappen. | |
| Herfra løber den slanke del, cauda, bagud. Den følger øvre flade af thalamus (8), hvor den ligger noget lateralt. | |
| Bøjer derefter nedad og siden fremad i temporallappen, hvor den ender ved corpus amygdaloideum (2). |
Side 326
| Nucleus caudatus (1) følger nøje lateralventriklen (7), som den er med til at begrænse. På grund af sin krumning ligger nucleus caudatus i bunden af lateralventriklen med sin øverste del, mens den nedadtil ligger i loftet af underhornet. | |
| Nucleus lentiformis (14) og (15) ligner en bikonveks linse med en stærkt konveks medialflade, og den er lige så lang som thalamus (12). Nucleus lentiformis ligger lateralt for thalamus og lateralt og under øverste del af nucleus caudatus (17). Nucleus lentiformis er skilt fra dem begge af den hvide substans i capsula interna (16). På frontalsnittet her er der tydelig adskillelse mellem nucleus caudatus (17) og nucleus lentiformis (14) og (15). | |
| Det svarer til et snit ved pilehoved (A). Et frontalsnit ved pilehoved (B) vil vise, at nucleus caudatus (1) og nucleus lentiformis (3) hænger sammen fortil. | |
| En betydelig del af thalamus (8) er skjult af nucleus lentiformis (3). De to strukturer er omtrent lige lange, men forskudt i forhold til hinanden. Thalamus når længere bagud end nucleus lentiformis. | |
| På snit gennem hjernen er nucleus lentiformis (14) og (15) let at identificere på grund af formen. Endvidere på grund af farveforskellen mellem det mediale lysere afsnit (15), “den blege kugle”, globus pallidus, og det mørkere laterale afsnit (14), “skallen”, putamen. Globus pallidus er bleg, fordi dens grå substans er blandet med hvid. | |
| Corpus amygdaloideum (2) ligger ved spidsen af nucleus caudatus i temporallappen. Corpus amygdaloideum er lille, i hvert fald sammenlignet med nucleus caudatus og nucleus lentiformis. Trods sin beskedne størrelse er corpus amygdaloideum en vigtig struktur, der omtales senere. | |
| Capsula interna er en vifteformet plade af hvid substans (4). Den smalle del af pladen (pile) er fortsættelsen af den hvide substans i crus cerebri i mesencephalon (17). På snit – horisontale og frontale – er capsula interna (16) et bredt bånd. Capsula interna ligger medialt for nucleus lentiformis (14) og (15) og lateralt for thalamus (12) samt øverste del af nucleus caudatus (17). Capsula interna indeholder vigtige ledningsbaner. | |
| Den almindelige form for hjerneblødning (1) er lokaliseret til capsula interna, og da motoriske baner ofte beskadiges af blødningen, indtræder halvsidig lammelse (2). |
| Ved agnosi forstås manglende evne til at fortolke sanseindtryk. En form for agnosi er “sjæleblindhed”. En velkendt genstand (1), mad skålen, kan ikke genkendes alene ved synets hjælp, selv om øje og synsbaner ikke fejler noget. Først ved berøring (2) identificeres skålen. |
Side 327
| En særlig form for sjæleblindhed ses (eksperimentelt og i klinikken) efter fjernelse af det meste af tindingelappen (3). Den viser sig ved at andre dyr (fx kat), som normalt afskyes af forsøgsdyret (hund), behandles på helt usædvanlig måde. Aber afskyer normalt slanger, men efter fjernelse af tindingelappen leger de gerne med dem. Ved afasi forstås cerebralt betingede forstyrrelser i sprogopfattelse og/eller sprogfrembringelse. | |
| Orddøvhed, sensitiv afasi, viser sig ved, at selv velkendte ord slet ikke opfattes (4), selv om hørelsen ikke fejler noget. Almindelig tale er lige så uforståelig som kinesisk for de fleste danskere. En anden form for afasi er ordblindhed, aleksi. Velkendte ord kan læses (5), men forståelsen svigter helt (6). Den almindelige (mildere) form for ordblindhed er dysleksi. | |
Side 328
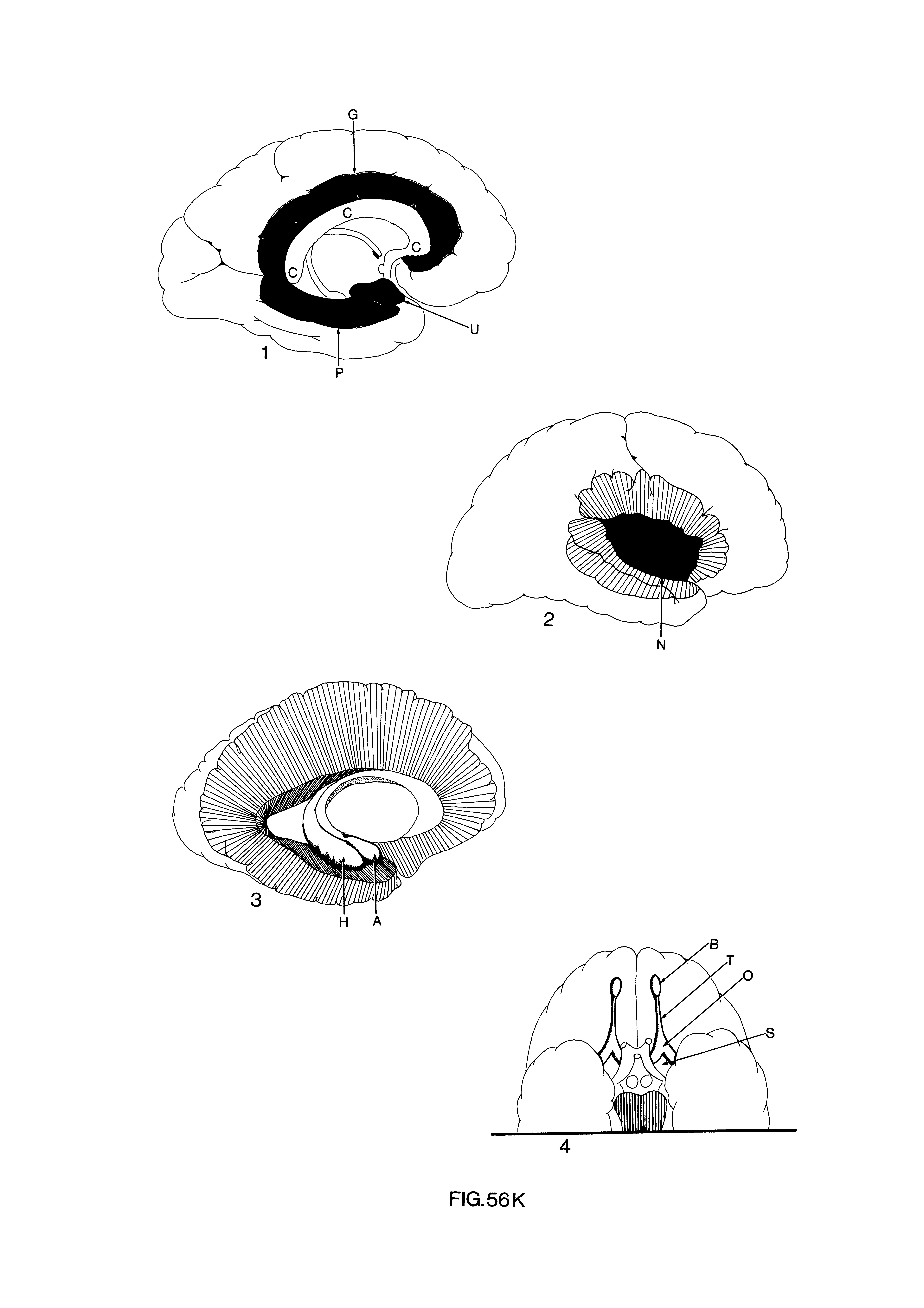
| Rhinencephalon, “lugtehjernen”, omfatter spredte afsnit af hemisphererne og svarer strengt taget kun til det område af hjernen, der modtager tråde fra lugtebanerne. Det limbiske system er derimod et godt eksempel på, at spredtliggende strukturer kan høre sammen i funktionel henseende, men netop fordi der ikke er tale om et samlet og funktionelt ensrettet system, anvendes mere og mere betegnelsen limbiske strukturer i stedet. Der er mange vigtige psykiske funktioner knyttet til de limbiske strukturer, og lugtesansen er kun en af dem. | |
| På basis cerebri findes dele, der udelukkende hører til rhinencephalon, og som ikke indgår i de limbiske strukturer. Her ligger “lugtekolben”, bulbus olfactorius (B). Den modtager lugtenerven, n. olfactorius, der er hjernenerve nr. 1, og løber fra lugteepitelet i næsehulen gennem sipladen til bulbus olfactorius. Fra lugtekolben (B) går impulserne bagud gennem et bundt nervetråde, tractus olfactorius (T), der er synlig på basis cerebri. Nervetrådene kan følges til området (O) lige foran substantia perforata anterior (S). | |
| De limbiske strukturer omfatter bl a | |
| hippocampus (H) i væggen af underhornet, | |
| fornix (2) mellem hippocampus og corpus mamillare, samt | |
| samt corpus amygdaloideum (A) ved underhornets spids. | |
| Desuden har dele af hypothalamus og thalamus tæt samarbejde med de limbiske strukturer og kan regnes med hertil. | |
Side 329
| Til de limbiske strukturer hører også hippocampus (H). | |
| Det er et fremspring, “en indre hjernevinding” (plusser), i den del af lateralventriklen (13), der ligger i tindingelappen og kaldes underhornet. Hippocampus har særlig betydning for indprentning af nye indtryk, og er en vigtig del af de limbiske strukturer. | |
| Fornix (2) er en ledningsbane. Den fører tråde (4) fra hippocampus (1) til corpus mamillare (3) i hypothalamus. Det er vigtigt for indprentningsevnen, at ledningsbaner fra hippocampus, især fornix, og selve hippocampus er intakte. Alkoholmisbrug kan fx medføre beskadigelser af hippocampus, med deraf følgende kraftigt nedsat evne til at huske “nye ting”. | |
| Til de limbiske strukturer hører endelig corpus amygdaloideum (2). Det er en mindre kerne ved spidsen af “halen” på nucleus caudatus (1), og det vil sige ved spidsen af underhornet, idet lateralventriklen (7) hele vejen ledsages af nucleus caudatus (1). Corpus amygdaloideum regnes både med til de limbiske strukturer og til basalganglierne. | |
| Sammenfattende kan man sige, at de limbiske strukturer (sort) ligger som et bælte af hjernevæv, der grænser op mod corpus callosum. Bæltet indeholder områder af cortex cerebri og kerneområder i storhjernen foruden dele af hypothalamus. Dele af dette område er hovedsæde for vores følelsesliv. Andre dele er væsentlige for hukommelsen. | |
| Forskellige følelser eller emotioner kan få os til at rødme, svede eller forandre puls. Alle disse reaktioner skyldes ændret aktivitet i det autonome nervesystem og hænger sammen med, at hypothalamus, der er det overordnede center for det autonome nervesystem, også indgår i de limbiske strukturer. | |
| Blandt sanserne indtager lugtesansen en særstilling, idet lugtenerven er koblet direkte til de limbiske strukturer. Denne sans er også meget effektiv til at fremkalde associationer af følelsesmæssig karakter. Der er få ting, der i højere grad end lugten af julebag og juletræ fører til oplevelsen af julestemning. | |
| De limbiske strukturer er “gamle hjerneafsnit” i udviklingsmæssig henseende. De kaldes undertiden den “viscerale hjerne”, fordi strukturerne spiller en rolle for mange af det autonome systems funktioner. Ved at stimulere limbiske strukturer, kan der fremkaldes forskellige autonome reaktioner. |
Side 330
| Det kan være vasomotoriske (1) (forsnævring af karrene til højre) eller ændring af tarmens peristaltik (2). Stimulation kan også fremkalde motoriske reaktioner bl a affektprægede som angreb eller forsvar (3) eller flugt (4). | |
| Funktionen af corpus amygdaloideum lader sig ikke beskrive med få ord. Ikke bare fordi vi stadigvæk forstår så lidt, men også fordi nyere forskning har vist en mangfoldighed af opgaver og forbindelser. Tendensen er dog tydelig. Amygdala er den del af hjernen, der har størst betydning for følelser og følelsesmæssig adfærd. Endvidere er det en hovedopgave, at etablere sammenhæng mellem et stimulus og dets følelsesmæssige værdi, altså om det, der sker, er “godt eller ondt”. Det er amygdala, der “ved”, at selvom “tante Mimi” taler venligt, er hun i virkeligheden “en led heks”. Den følelsesmæssige farvning af andres tale opfanges af amygdala. | |
| Hippocampus er vigtig for indprentning af nye indtryk. Ved læsion af hippocampus, fornix eller corpus mamillare er patienten ude af stand til at huske, hvad han/hun lige har læst eller hørt. Fjernelse af en tindingelap medfører en form for “sjæleblindhed”, idet dyr, der normalt ikke tolereres af forsøgsdyret, nu behandles “venligt” (3). | |
| På aber har fjernelse af begge tindingelapper medført, at aggressive dyr bliver særdeles fredelige, samtidig med at de får en enorm appetit både på føde og på et intenst seksualliv. Desuden bliver hukommelsen dårlig, da hippocampus fjernes sammen med resten af temporallappen. | |
| De to lateralventrikler (1) er U-formede med konkaviteten fremad. Gennem foramen interventriculare står de i forbindelse med tredie ventrikel (2). | |
| Foramen interventriculare (5) har en beskeden størrelse. Det ligger i lateralvæggen af tredie ventrikel ved forreste ende af thalamus (T). |
Side 331
| Hver lateralventrikel består af tre horn og en central del. Der er et forhorn (1) i lobus frontalis, et baghorn (2) i lobus occipitalis og et | |
| underhorn (3) i lobus temporalis. Den centrale del (4) forbinder de tre horn. Den (13) ligger på øvre flade af thalamus (12) under corpus callosum (C). | |
| I medialvæggen af både den centrale del (øverste 13) af lateralventriklen og underhornet (13) findes tela choroidea (T). Tela choroidea danner liquor cerebrospinalis. | |
| Forhorn (1) og baghorn (2) mangler tela choroidea, mens den centrale del og underhornet har en sammenhængende stribe tela choroidea (5) i medialvæggen. | |
| Fjerde ventrikel har også tela choroidea i loftet, men kun i nederste del (2) under cerebellum (3). Tela choroidea består overalt af et lag ependymceller (7) og pia mater (skraveret). Liquor cerebrospinalis produceres i alle afsnit af tela choroidea og løber til fjerde ventrikel. Gennem hullerne nedadtil i loftet af fjerde ventrikel løber væsken | |
| ud i spatium subarachnoideum (5) omkring centralnervesystemet. Herfra strømmer væsken over i blodbanen, sinus durae matris (10), gennem udposninger (9) af arachnoidea (4) i dura mater. | |
| En ledningsbane er en neuronkæde (1), (5) og (9), der kan lede impulser. Neuronerne kobles sammen af synapser, der ligger i den grå substans (6 )og (8) i centralervesystemet. Nervetrådene (3), (7) og (10) i ledningsbanerne, og det vil sige neuronernes lange udløbere, ligger i den hvide substans. | |
| De vigtigste banetyper er commissurbaner (1) mellem højre og venstre halvdel af hjerne eller rygmarv, associationsbaner (2) mellem rygmarvssegmenter i samme side eller mellem hjerneafsnit i samme side og projektionsbaner (3) mellem hjerne og rygmarv. Når impulserne går mod hjernen, er banen afferent, og når impulserne går fra hjernen, er banen efferent |
Side 332
| Commissurbaner i rygmarven løber gennem den hvide substans lige bag ved fissura mediana anterior. Associationsbaner i rygmarven danner en smal bræmme (mørkt prikket) nærmest den grå substans i bagstreng (3), side- og forstreng (1). Projektionsbaner optager resten af den hvide substans i rygmarven. I bagstrengen er alle projektionsbaner afferente (1) og (2), mens der i sidestrengen er både afferente (2‑4) og efferente (5‑6). Det samme gælder i forstrengen med både afferente (2) og efferente (3) og (4). |
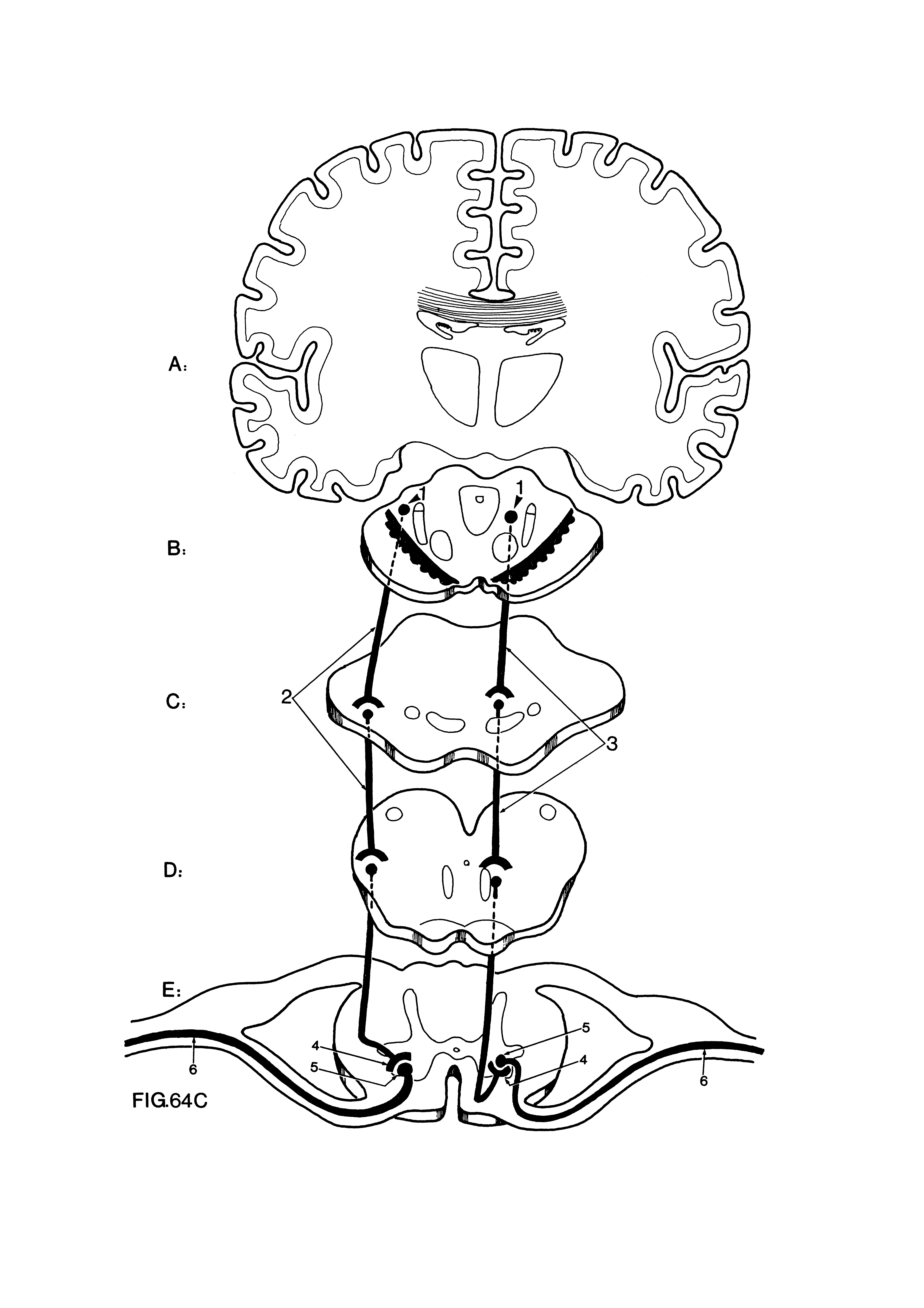
| Afferente eller sensitive ledningsbaner omfatter primære og sekundære baner. | |
| En primær ledningsbane er kun repræsenteret af første neuron (1‑3) i medulla spinalis, dvs, at synapsen mellem første og andet neuron ligger helt oppe i hjernestammen (6). Alle primære baner ligger i bagstrengen i medulla spinalis og er derfor identiske med bagstrengsbaner. En sekundær ledningsbane har synapsen mellem første og andet neuron liggende i baghornet (6). |
| Første neuron (1), (2) og (3) i en primær sensitiv ledningsbane er en pseudounipolar nervecelle, hvis cellelegeme (1), dvs neuronets trofisk centrum, ligger i et spinalganglion. Dendriten (2) fører impulser fra periferien, og de fortsætter gennem neuriten (3), der løber til medulla spinalis gennem radix posterior (4). | |
| Første neuron danner ikke synapse i medulla spinalis, men neuriten ascenderer i bagstrengen til medulla oblongata (snit D), hvor den danner synapse med andet neuron (5) i en bagstrengskerne (6).
Da vi kun træffer første neuron i medulla spinalis, kaldes banen for primær, og første neuron strækker sig altså fra periferien til en bagstrengskerne i medulla oblongata. |
|
| Andet neuron er en multipolar nervecelle (5,7), der strækker sig fra en bagstrengskerne (6 eller 6a) til thalamus (8) i modsat side. Cellelegemet (5), og dermed trofisk centrum for neuronet, ligger i en bagstrengskerne (6), og det er neuriten (7), der krydser midtlinien. |
Side 333
| Tredie neuron er en multipolar nervecelle (9) og (10), der strækker sig fra thalamus (8) til cortex cerebri, gyrus postcentralis (11). Trofisk centrum for tredie neuron er cellelegemet (9) i thalamus (8). | |
| I bagstrengen i medulla spinalis ordner bagstrengsbanerne sig i to grupper. En gruppe medialt (1), der fører impulser fra underekstremitet og nederste del af kroppen, og en gruppe lateralt (2), der fører impulser fra overekstremitet og øverste del af kroppen. | |
| De to grupper går til hver sin bagstrengskerne i medulla oblongata, (6) og (6a), der indeholder trofisk centrum for andet neuron. Neuriterne herfra krydser midtlinien i (D). Efter krydsningen fortsætter de i et samlet bundt (L) op gennem hjernestammen til thalamus. |
| Bagstrengsbanerne fører “dybdesensibilitet”, dvs proprioceptive impulser fra muskler, sener og led samt tryk- og berøringsindtryk fra huden på krop og ekstremiteter. Patienter med læsion af bagstrengsbanerne kan stå normalt, når de kan se, men lukker de øjnene, falder de, idet de mangler orienteringen om kroppens stilling i rummet på grund af den manglende dybdesensibilitet. Gangen er “spjættende” og usikker. Det gælder især, når de ikke kontrollerer benenes stilling med synet. | |
| Synapsen mellem første og andet neuron i en sekundær ledningsbane ligger i rygmarvens baghorn, dvs, andet neuron er repræsenteret i rygmarven i modsætning til primære sensitive ledningsbaner, hvor det kun er første neuron, der findes i rygmarven. |
Side 334
| Sekundære sensitive ledningsbaner findes i rygmarvens sidestreng og forstreng. | |
|
I bagstrengen findes ingen sekundære sensitive ledningsbaner kun primære, og de primære er udelukkende placeret her. |
Side 335
| En ledningsbane fra rygmarven til thalamus (lateralt) fører smerte- og temperaturindtryk, mens en anden (fortil) fører tryk- og berøringsindtryk. | |
| Ledningsbanerne fra rygmarven til lillehjernen (1a) og (1b) går til paleocerebellum (2). De fører dybdesensibilitet, dvs proprioceptive impulser. Derved orienteres cerebellum om musklernes |
Side 336
| spændingstilstand, leddenes stilling mv, uden at denne orientering bliver bevidst. Det er bl a på grundlag af oplysningerne gennem disse ledningsbaner, at cerebellum kan foretage justeringer og korrektioner af bevægelser, der planlægges af storhjernen. |
| En ledningsbane føre nerveimpulser fra periferien til formatio reticularis (10), (11) og (12) i hjernestammen. Funktion omtales under beskrivelsen af formatio reticularis. |
| Efferente eller motoriske rygmarvsbaner fører impulser fra hjernen til rygmarvens forhorn. Der er to hovedtyper af forbindelser mellem de motoriske centre i hjernen og de motoriske forhornsceller i rygmarven. | |
| Forbindelsen kan være direkte uden synapser undervejs. Disse neuriter fra cortex cerebri ender i medulla spinalis med en synapse direkte på en forhornscelle eller på et indskudsneuron, som er koblet til forhornscellen. Den type ledningsbaner fra cortex cerebri kaldes pyramidebaner. | |
| De øvrige forbindelser til den motoriske forhornscelle kaldes for de ekstrapyramidale baner, hvor neuriten kommer fra nerveceller i kerneområder i hjernestammen. Neuriter fra cerebellum kan påvirke disse subcorticale kerneområder, eksempelvis nucleus ruber, substantia nigra, formatio reticularis og vestibularkerner, og derved ændre på impulserne til den motoriske forhornscelle | |
| Banerne med impulser til forhornene er de bedst kendte. Der findes også baner til lateralhornene. De indgår i den autonome del af nervesystemet. | |
Side 337
| I praksis er der ingen skarp adskillelse af de opgaver, der varetages af pyramidebanerne og de, der varetages af de ekstrapyramidale baner. Systemerne samarbejder under udførelsen af forskellige bevægelser, herunder ændringer af balance- og kropsstilling, når det er nødvendigt. |
| Pyramidesidestrengsbanen, består af to lange neuroner. Første neuron strækker sig fra hjernebark (2) til forhorn (8), andet neuron fra forhorn (8) til periferi (muskel). | |
| Første neuron har trofisk centrum i den motoriske del af hjernebarken, gyrus præcentralis (2). Nervecellelegemet er pyramideformet og ikke stjerneformet som i almindelige multipolare nerveceller. Neuriten løber gennem capsula interna (pil) og fortsætter til hjernestammen. Her løber den gennem mesencephalon (snit B), hvor den ligger i crus cerebri. Derefter gennem pons (snit C), hvor den ligger fortil, og til medulla oblongata (snit D). | |
| Nedadtil i medulla oblongata krydser neuriten midlinien i (5). Den fortsætter i medulla spinalis (snit E), hvor den løber ned gennem sidestrengen (7) til en forhornscelle, dvs en multipolar nervecelle, hvis cellelegeme (9) ligger i forhornet (8). Forhornscellen er trofisk centrum for andet neuron, og neuriten (10) herfra forlader medulla spinalis gennem radix anterior (11) og løber videre i spinalnerven (12). | |
| Pyramideforstrengsbanen består af to lange neuroner. Første neuron har igen trofisk centrum (1) i gyrus præcentralis, og neuriten løber gennem capsula interna (pil) og hjernestammen. Modsat pyramidesidestrengsbanen sker der ingen krydsning i medulla oblongata. Tværtimod fortsætter neu- |
Side 338
| riten (6) ukrydset til medulla spinalis, hvor den løber i forstrengen (13). Midtlinien (14) krydses først i den højde, hvor andet neuron har trofisk centrum (9), og højden afhænger af, hvilken tværstribet skeletmuskel det pågældende neuron skal innervere. | |
| Synapsen mellem første og andet neuron findes i forhornet (8), og andet neurons neurit (10) løber perifert gennem radix anterior (11). Pyramideforstrengsbanen udgør kun ¼ af de pyramidebaner, der går til rygmarven, og de to vigtigste forskelle på denne bane og pyramidesidestrengsbanen er altså beliggenheden af første neurons krydsning og banens placering i rygmarven. | |
| Pyramideforstrengsbanen krydser i medulla spinalis, og indtil krydsningen løber de i forstrengen (13). Pyramidesidestrengsbanen krydser i (5) i medulla oblongata. Efter krydsningen løber de i sidestrengen (7) i medulla spinalis. |
| Pyramidebanerne i for- og sidestreng sender impulser til tværstribet muskulatur innerveret af rygmarvsnerver. |
| Ved beskadigelse af første neuron (1) i pyramidebanerne fås en central eller supranucleær lammelse. Årsagen er ofte blødning i capsula interna. En beskadigelse af andet neuron resulterer i en perifer lammelse. Lammelsen kaldes nucleær, hvis det er cellelegemerne (2) i forhornet, der er beskadiget, og infranucleær, hvis det er forhornscellernes neuriter (3), der er læderet. En nucleær lammelse ses ved polio, og en infranucleær lammelse ses ved perifere nervelæsioner. | |
| Karakteristisk for en perifer lammelse er mangel på reflekser, arefleksi, bortfald af tonus, atoni og efterhånden svind af muskulaturen, atrofi. Hovedsymptomet er naturligvis lammelse. | |
| Årsagen til arefleksi er læsionen af andet neuron, der fungerer som det efferente neuron i en refleksbue. Muskeltonus opretholdes gennem reflekser. Derfor er det klart, at atoni indtræder, når refleksbuernes efferente neuron ødelægges. Når muskler mangler nerveforsyning indtræder inaktivitetsatrofi, dvs, muskulaturen svinder, fordi den ikke kan bruges. Ved en central lammelse, der hyppigst skyldes blødning i capsula interna, er reflekserne livlige, tonus udtalt og der mærkes en vis |
Side 339
| “stivhed” i muskulaturen, spasticitet, der vanskeliggør passive bevægelser i leddene. Det vil sige, at det fx er vanskeligt for undersøgeren at bøje patientens arm. | |
| De forskellige symptomer skyldes at refleksbuerne er intakte, og at formatio reticularis, med sin overvejende fremmende virkning på tonus, ikke længere hæmmes fra højereliggende områder. | |
| Muskelstivheden, spasticiteten, er meget karakteristisk og skyldes øget proprioceptiv refleksvirksomhed. Når undersøgeren bøjer patientens arm i albuen, strækkes musklerne på bagsiden af armen. | |
| Fra muskeltenene her udløses afferente impulser til forhornsceller, som innerverer musklerne på bagsiden af armen. Musklerne kontraherer sig, og da de fungerer som strækkemuskler for albueleddet, modvirker de bøjebevægelsen. Armen føles derfor unaturlig stiv. De samme reflekser udløses også normalt, men her dæmpes de af hjernen, og derfor kan armen hos en rask person bøjes uden modstand. |
Side 340
| Selv om pyramidebanerne er meget vigtige fx for aktivering af muskelgrupper i forbindelse med bevægelser, der kræver stor opmærksomhed, så har det ekstrapyramidale system stor betydning for aktivering af muskelgrupper, hvor et vel afstemt samspil er nødvendigt for stabil kropsstilling og balance eller glidende bevægelser, fx gang. | |
| Den motorisk forhornscelle (5) “bombarderes” således med impulser fra både hjernebark gennem pyramidebanerne (1) og fra subcorticale kerner hørende til det ekstrapyramidale system gennem forskellige baner. De enkelte signaler til forhornscellen kan være fremmende eller hæmmende, og det er det samlede resultat, der bestemmer styrken af de signaler, forhornscellen videresender til de muskelceller, den innerverer. |
| Det er vigtigt at fastslå, at ledningsbanerne med forbindelse til hjernenerverne principielt løber og fungerer på helt samme måde, som ledningsbanerne med forbindelse til rygmarvsnerverne. | |
| Hjernenerverne har apparent udspring fra hjernestammen, og banernes forløb udenfor denne del af centralnervesystemet sker gennem nerver, vi kalder hjernenerver, mens forløbet af nervebanerne udenfor rygmarven sker gennem rygmarvsnerver. | |
| I begge tilfælde vil de efferente ledningsbaner til de motoriske endeplader i den tværstribede muskulatur omfatte to neuroner, hvor andet neuron enten har sit trofiske centrum i en motorisk hjernenervekerne i hjernestammen eller i en motorisk forhornscelle i medulla spinalis og herfra fortsætte til effektororganet, den motoriske endeplade. Og i begge tilfælde gælder definitionen på en motor unit, som er en motorisk nervecelle med alle de tværstribede muskeltråde, den innerverer. I muskler med betydning for fine og præcise bevægelser er der langt færre muskeltråde pr nervecelle end i eksempelvis sædemusklen |
Side 341
| På samme måde er forløbet af de afferente nervetråde opbygget identisk. Dendriten fra en pseudounipolar nervecelle, eller i sjældne tilfælde en bipolar, fører impulsen fra receptororganet i periferien i retning mod centralnervesystemet. Nervecellelegemet for dette neuron, som er det første i ledningsbanen, ligger i et sensitivt ganglie. | |
| Når det drejer sig om de afferente nervetråde i rygmarvsnerverne, ligger den pseudounipolare nervecelle, dvs det trofiske centrum for første neuron, i et spinalganglie. Herfra fortsætter neuriten gennem radix posterior til medulla spinalis. | |
| Når det drejer sig om de afferente nervetråde i hjernenerverne, begynder det første neuron igen med at lede impulser fra receptororganet gennem dendriten og ind til et sensitivt hjernenerveganglie, der sædvanligvis består af pseudounipolare nerveceller nøjagtig som ved spinalganglierne. | |
| Fra det sensitive hjernenerveganglie, der samtidig er trofisk centrum for første neuron i den sensitive hjernenervebane, fortsætter neuriten ind i hjernestammen, hvor den ender med at danne synapse i en terminalkerne. |
| Første neuron (1,2,3) i en sensitiv hjernenervebane er som hovedregel en pseudounipolar nervecelle. Trofisk centrum, dvs cellelegemet (1), ligger i et sensitivt hjernenerveganglion. Dendriten (2) modtager fx sensitive indtryk fra huden på ansigtet eller slimhinden i mundhulen, og neuriten (3) fortsætter gennem hjernenerven til en sensitiv hjernenervekerne (4) i hjernestammen, hvor den danner synapse med en multipolar nervecelle. | |
| Neuriten fra andet neuron (5) i den sensitive hjernenervekerne krydser midtlinien og ascenderer i hjernestammen til thalamus, hvor den danner synapse med tredie neuron (6), en multipolar nervecelle, hvis neurit (7) går til hjernebarken, gyrus postcentralis (8). Ligheden med de sensitive rygmarvsbaner må siges at være slående. | |
| Når alle de forskellige sensitive ledningsbaner løber op gennem hjernestammen, vil banerne, der dannes af andet neuron, danne et tydeligt bundt eller bånd. | |
| Da hjernenerve nr. 5, n. trigeminus, er følenerve for hele ansigtet, indeholder den så mange sensitive nervetråde, at forløbet af andet neuron fra de sensitive trigeminuskerner til thalamus danner et bundt. |
Side 342
| Når vi har omtalt pyramidebaner indtil nu, har vi kun beskæftiget os med de motoriske pyramidebaner til medulla spinalis. I virkeligheden omfatter pyramidebaner efferente eller motoriske baner til både medulla spinalis og til motoriske hjernenervekerner i hjernestammen. | |
| Da pyramidebanerne til medulla spinalis omfatter langt de fleste nervetråde, bruges betegnelsen “pyramidebane” ofte om dem alene, og det er forkert. De pyramidebaner, der går til hjernenervekerner, hører også med. | |
| Første neuron i alle pyramidebaner er en multipolar nervecelle (9) med et pyramideformet cellelegeme i den motoriske zone af cortex cerebri. Neuriten (10) går gennem capsula interna til hjernestammen, og når den skal til en motorisk hjernenervekerne (11), ender den i hjernestammen, idet den danner synapse med en multipolar nervecelle i den pågældende kerne (11). De fleste aksoner (10) krydser midtlinien inden synapsen. | |
| Neuriter, der danner synapse i oculomotorius eller trochleariskernen, ender naturligvis i mesencephalon (B), mens neuriter, der skal til trigeminus- eller facialiskernen, først ender i pons (C). Andet neuron har trofisk centrum i den motoriske kerne (11), der fungerer som forhorn, og neuriten (12) løber perifert som en tråd i den hjernenerve, den hører til. |
| Ved en refleks forstås den reaktion, der finder sted, når afferente impulser (via 5, 4, 6), gennem en eller flere synapser (8) i centralorganet, udløser efferente impulser (via 9), der fremkalder motorisk (1) eller sekretorisk aktivitet. | |
| En simpel refleksbue består af to neuroner, et afferent og et efferent. Trofisk centrum for det afferente neuron er cellelegemet af en pseudounipolar nervecelle (4), og trofisk centrum for det efferente neuron er cellelegemet af en multipolar nervecelle i et forhorn (8). De to neuroner danner synapse i forhornet. Som det fremgår af definitionen, kan flere synapser være inddraget i en refleks, og i komplicerede refleksbuer er antallet af synapser stort. Vi skelner mellem proprioceptive eller dybe reflekser og eksteroceptive eller overfladereflekser. |
Side 343
| Proprioceptive eller dybe reflekser kan benytte en simpel refleksbue i medulla spinalis bestående af et afferent og et efferent neuron. Trofisk centrum for det afferente neuron er cellelegemet af en pseudounipolar nervecelle (4) i et spinalganglie. Dendriten (5) starter i en muskelten (2) og løber gennem spinalnerven (11). Neuriten (6) går gennem radix posterior (7) til medulla spinalis. Her danner den synapse med en forhornscelle (8). | |
| Neuriten (9) fra den multipolare forhornscelle går gennem radix anterior (10), og spinalnerve (11) til musklen (1). Ved strækning af musklen strækkes muskeltenen (2), og derved udløses impulser gennem det afferente neuron (5, 4, 6). Impulserne overføres til det efferente neuron (9), og resultatet bliver en kontraktion af musklen (1). | |
| Det er en refleksbue af denne type, der sættes i funktion, når knærefleksen fremkaldes. Det sker ved, at man slår på ligamentet mellem knæskal og skinneben. | |
| Proprioceptive reflekser er grundlaget for musklernes “permanente spændingstilstand”, også kaldet muskeltonus. Muskeltonus skyldes altså en permanent refleksaktivitet, der holdes i gang af impulser fra formatio reticularis gennem motoriske gammanervetråde til muskeltene. | |
| En muskelten rummer flere tynde, intrafusale muskelceller (4). Kun enderne af cellerne har tværstribning, og det er kun de tværstribede ender af cellen, der er i stand til at trække sig sammen. Det sker, når de får impulser gennem motoriske tråde, gammatråde (2). | |
| Når midterstykket strækkes, enten ved at enderne af de intrafusale muskelceller trækker sig sammen, eller ved at hele den omgivende tværstribede muskel strækkes, udløses der nerveimpulser gennem den sensitive tråd (3), der danner spiral om midterstykket, og som er det afferente neuron i refleksbuen. | |
| Afgørende for udløsning af afferente impulser – og dermed reflekser – er strækningen af det ikke-tværstribede midterparti i de små intrafusale muskelceller. Ved knærefleksen strækkes midterpartiet af muskeltenens intrafusale muskelceller sammen med hele musklen på grund af slaget mod ligamentet. |
Side 344
| Ved tonus strækkes midterpartiet på grund af de tværstribede enders kontraktion, og kontraktionen skyldes impulser fra formatio reticularis mv. |
| Eksteroceptive reflekser benytter refleksbuer, hvor det afferente neurons dendrit begynder i hud eller underhud enten som frie nerveender eller ved specielle endeorganer. Kradsningsrefleksen hos hunden og plantarrefleksen hos mennesket er eksempler på eksteroceptive reflekser. Kradsningsrefleksen fremkaldes ved at klø en hund på den ene side af kroppen, og refleksen viser sig ved rytmiske bevægelser af bagbenet i samme side. Plantarrefleksen viser sig ved at tæerne bøjes (mod underlaget), når man stryger på en bestemt måde i fodsålen. | |
| Det ekstrapyramidale system er en vigtig del af det motoriske system. Det består af spredte strukturer i telencephalon, hjernestamme og cerebellum. Alle strukturerne er allerede omtalt mere detaljeret. I dette afsnit fokuseres på, hvorledes disse spredte kerneområder har stor betydning for det samlede motoriske system. | |
| Det ekstrapyramidale system har vigtige udfaldsveje. De går dels til medulla spinalis, hvor de danner synapse med forhornsceller, dels til motoriske hjernenervekerner, hvor de ligeledes danner synapse med multipolare nerveceller. | |
| Basale hjerneganglier (1) står i forbindelse med cortex cerebri (10). |
Side 345
| Endvidere er der mange indbyrdes forbindelser mellem komponenterne i det ekstrapyramidale system. Ovenikøbet er forbindelserne som regel reciprokke, dvs løber begge veje. | |
| Spredte strukturer danner et system, når de bidrager til de samme funktioner. Det er tilfældet for komponenterne i det ekstrapyramidale system, og betydningen af systemet kan bedst vises, ved at beskrive symptomer, der optræder ved sygdomme i det ekstrapyramidale system. Hovedsymptomerne er ufrivillig bevægelse og rigiditet. De ufrivillige bevægelser kan være forskellige. Bedst kendt er en regelmæssig fingertremor (tremor = rysten), der benævnes “pilletrillertremor” (6). Den ses især hos ældre mennesker, men kan også ramme yngre. | |
| Rigiditet betyder stivhed, og her tænkes på den tonusforøgelse i muskulaturen, som ses ved lidelser i det ekstrapyramidale system. I modsætning til spasticitet, der bl a ses ved hjerneblødning, er rigiditet mindre tilbøjelig til at “give sig”. | |
| Den viser sig ved en sej vedvarende modstand mod passive bevægelser (3). Det er næsten ikke muligt, at rette armene ud eller bøje dem. Tonusforøgelsen kan være så voldsom i hele kroppen, at patienten bogstavelig talt bliver stiv som en statue, statuemenneske. | |
| Ejendommelig nok optræder både ufrivillige bevægelser og rigiditet ofte samtidig. Resultatet bliver, at patienten går stift og let foroverbøjet (2) samtidig med, at der er tremor i underarme, hånd og fingre, Parkinsons sygdom. |
| Formatio reticularis er en del af det ekstrapyramidale system, men omtales specielt på grund af dens betydelige udstrækning og store funktionelle betydning. Formatio reticularis (4) findes gennem hele hjernestammen og består af mange små øer af kerner, der modtager impulser fra næsten alle dele af centralnervesystemet samt fra afferente ledningsbaner. Formatio reticularis sender både impulser opad til højere liggende |
Side 346
| hjerneafsnit og nedad til lavere liggende afsnit af centralnervesystemet. Formatio reticularis har dermed to hovedfunktioner, en ascenderende og en descenderende. |
| Den ascenderende funktion består i, at formatio reticularis (4) både påvirker hjernebarken (1) direkte og indirekte gennem thalamus (2) og hypothalamus (5). Denne påvirkning har betydning for bevidsthedsniveauet. Så længe hjernebarken “bombarderes” med impulser fra formatio reticularis, vil impulserne gennem de sensitive ledningsbaner (3) registreres i hjernebarken. Hvis formatio reticularis ophører med at sende impulser til hjernebarken, vil signaler gennem de sensitive ledningsbaner, uanset signalernes styrke, ikke blive modtaget, og patienten er bevidstløs. | |
| Når aktiviteten i formatio reticularis falder til et vist niveau, fx ved træthed, falder man i søvn (7). | |
| Formatio reticularis (2) har en descenderende funktion af stor betydning for muskeltonus. En større del af formatio reticularis virker fremmende og en mindre del hæmmende på tonus. Begge dele er under påvirkning af impulser fra bl a cortex cerebri (1), der virker regulerende på formatio reticularis. Hvis forbindelsen med cortex (4) afbrydes, løber formatio reticularis “løbsk” med en stærk tonusforøgelse (5) som resultat. | |
| Muskeltonus forudsætter permanent refleksvirksomhed, og det er formatio reticularis, der sender impulser til forhornsceller (9), hvorfra gammamotoriske tråde (2), går til de små intrafusale muskelceller (4) i muskeltenene. De er kun kontraktile i enderne, hvor der er tværstribning, ikke i midten hvor sensitive nervetråde (3) danner |
Side 347
| spiral og registrerer strækning af muskelcellens midterparti, når endepartierne trækker sig sammen. Den sensitive nervetråd (3) leder impulser til store multipolare nerveceller i forhorn. Herfra fører alfamotoriske tråde (5) impulser til tværstribede muskelceller (6), hvor der sker kontraktion. |
| Ved total overskæring (2) af medulla spinalis lige under foramen magnum er forbindelsen mellem hjerne og rygmarv helt afbrudt, og resultatet kaldes et spinaldyr (5). | |
| Spinaldyret ligger slapt hen, fordi tonus i musklerne er faldet bort. Forudsætningen for muskeltonus er, at formatio reticularis vedligeholder den konstante, svage refleksvirksomhed. De dybe reflekser kan dog let fremkaldes og er livlige, fordi hjernen ikke kan hæmme dem. Så snart dyret overlades til sig selv, dør reflekserne imidlertid hurtigt hen. De er ikke permanente, fordi formatio reticularis ikke kan sende impulser til forhornene på grund af snittet (2) gennem rygmarven. | |
Side 348
| For at vise hvordan formatio reticularis påvirkes af andre hjerneafsnit, har neurofysiologer skåret hjernestammen igennem i forskellige niveauer. |
| En overskæring ved øvre kant af pons (1) kaldes decerebrering. Blodtrykket (2) opretholdes på et nogenlunde normalt niveau, men muskeltonus er stærkt forøget, og tilstanden kaldes decerebreringsrigiditet (3). | |
| Årsagen er hyperaktivitet i de proprioceptive refleksbuer (3‑5), fordi hjernebarken ikke mere bremser formatio reticularis, hvis fremmende virkning derfor slår igennem med fuld kraft i rygmarven. | |
| Tonusfordelingen i musklerne, særlig i ekstremiteterne, påvirkes af halsmusklerne og af hovedets stilling. De halsreflekser, der har betydning for tonusfordelingen, kaldes toniske, og de udløses fra halsens muskler og led. Toniske halsreflekser er med andre ord reflektorisk udløste ændringer af tonusfordelingen i kroppens tværstribede muskulatur, og reflekserne kommer fra receptorer omkring led og muskler i halsens dybe dele. | |
| De toniske hals- eller nakkereflekser bidrager til, at kroppens stilling i forhold til hovedet holdes konstant. Her kan indskydes, at labyrintreflekser bidrager til at holde hovedets stilling i rummet konstant. | |
Side 349
| Hos et midthjernedyr skæres hjernestammen igennem ved øvre kant af mesencephalon (eller et lille stykke ind i diencephalon), dvs, hele hjernestammen og cerebellum er intakt. Midthjernedyret ligner det decerebrerede dyr i motorisk henseende. Det står omtrent på samme måde som det decerebrerede dyr, men mere naturligt. Det adskiller sig dog på afgørende måde fra det decerebrerede dyr ved at kunne rejse sig igen, når det væltes. | |
Side 350
| Det perifere nervesystem består af alle nerveceller, nervetråde og tilhørende gliaceller, fx Schwanns’ celler, uden for centralnervesystemet. Al information til og fra centralnervesystemet går via 12 par hjernenerver og 31 par spinalnerver. | |
| Det er naturligt, at inddele det perifere nervesystem i en sensorisk, afferent del, som sender informationer ind til centralnervesystemet, og en motorisk, efferent del, som sender information fra centralnervesystemet til muskler og kirtler, dvs effektorer. | |
| Ved gennemgang af en perifer nerve, hjernenerve eller rygmarvsnerve, er vi interesseret i nervens apparente udspring, dvs det sted på overfladen af hjerne eller rygmarv, hvor nerven kommer til syne, perforation af dura mater og det hul, som nerven benytter, når den forlader kraniehulen eller kanalen i hvirvelsøjlen. | |
| Udtrykket “trofisk centrum” bruges om det sted, hvor nervecellelegemerne findes for de nervetråde, der viser sig i det apparente udspring. I stedet for “trofisk centrum” bruges undertiden udtrykket nervetrådenes reelle eller “virkelige” udspring. | |
Side 351
| Konklusionen er, at trofisk centrum for sensitive nervetråde i det apparente udspring findes i ganglier uden for centralnervesystemet, mens trofisk centrum for motoriske tråde i det apparente udspring ligger i centralnervesystemet. |
| Man skelner fra gammel tid mellem 12 par hjernenerver, som nummereres efter den rækkefølge, de perforerer dura i kraniebunden. Nummereringen begynder fortil og fortsætter i retning bagud. Ved at se på hullerne i basis cranii interna, kan man nogenlunde regne ud, hvor nerverne perforerer dura. |
| Oversigten kan i første omgang bruges til orientering. Når afsnittet om hjernenerverne er læst, kan hovedpunkter opfriskes ved hjælp af oversigten. |
| Nr. | navn | fig. | hovedfunktion |
|---|---|---|---|
| 1 | n. olfactorius | 1&2 | lugtenerve |
| 2 | n. opticus | 3 | synsnerve |
| 3 | n. oculomotorius | 5 | bevægenerve (1) for øjeæble m. rectus medialis, superior, inferior, m. obliquus inferior, øvre øjelåg m. levator palpebrae sup. parasympatiske tråde via ggl. ciliare til akkomodationsmusklen (8) og pupillens ringmuskel (7). |
| 4 | n. trochlearis | 6 | bevægenerve (2) for øjeæble m. obliquus superior. |
Side 352
| Nr. | navn | fig. | hovedfunktion |
|---|---|---|---|
| 5 | n. trigeminus | 7 | ansigtets, øjenhulens, næsehulens og mundhulens følenerve (6), (7), (8) bevægenerve, se 53 |
| 51 | n. ophthalmicus, første trigeminus gren | 7 | følenerve for øjenhule, pande-og en del af næsehule (6) |
| 52 | n. maxillaris, anden trigeminusgren | 20 & 7 | følenerve for overkæbe, inkl. tænder og tandkød, ansigt og en del af næsehule (7) |
| 53 | n. mandibularis, tredie trigeminusgren | 21 & 7 | følenerve for underkæbe, inkl. tænder og tandkød, tungeryggens forreste del og kind mv (8). bevægenerve for tyggemuskler, mundbundsmuskler, én ganemuskel og én muskel i trommehulen. |
| 6 | n. abducens | 6 | bevægenerve (3) for øjeæble, m. rectus lateralis. |
| 7 | n. facialis | 22 & 23 | ansigtets bevægenerve, bevægenerve for ansigtsmuskler (mimiske muskler), platysma, venter posterior m. digastrici, m. stylohyoideus og m. stapedius.
parasympatiske, sekretoriske tråde til tårekirtel (7), næsekirtler (8) og ganekirtler (9) via ggl. pterygopalatinum (12). Og til gl. submandibularis (10) og gl. sublingualis (11) via ggl. submandibulare (13), smagstråde fra forreste del af tungeryggen. |
| 8 | n. vestibulocochlearis | 4 | ligevægts- og hørenerve (1) og (2) |
Side 353
| Nr. | navn | fig. | hovedfunktion |
|---|---|---|---|
| 9 | n. glossopharyngeus | 24 & 25 | følenerve for tungeryggens bageste halvdel (7), mandler (8) og svælg. Vigtigste smagsnerve og registrerer blodtryk og blodets iltindhold (5). Bevægenerve for én svælgmuskel, m. stylopharyngeus (6), parasympatiske (sekretoriske) tråde til gl. parotidea (10) via ggl. oticum (11). |
| 10 | n. vagus | 27 | bevægenerve for gane (8) undtagen m. tensor veli palatini, svælg (9) undtagen m. stylopharyngeus, strubehoved (10) og spiserør (6) parasympatiske tråde til fordøjelseskanal fra tunge til flexura coli sinister, bugorganer, pancreas (17), milt (15), lever (13) og galdeblære, brystorganer, luftveje (12) og hjerte (11), følenerve for de samme områder, få smagstråde |
| 11 | n. accessorius | 29 & 30 | hjælpenerve for n. vagus med motoriske tråde bevægenerve for m. sternocleidomastoideus (6) og m. trapezius (7) |
| 12 | n. hypoglossus | 31 | bevægenerve (1) for tungen undtagen musklen til gane, m. palatoglossus og undtagen musklen til svælget, pars glossopharyngea af m. constrictor pharyngis superior |
Side 354
| De tre nerver er “sansenerver” eller sensitive nerver og beskrives under ét. Udtrykket sansenerve bruges ofte om de nerver, der har med de “klassiske” sanser at gøre. N. olfactorius, hjernenerve nr. 1, er lugtenerven. N. opticus, hjernenerve nr. 2, er synsnerven, og n. vestibulocochlearis eller n. statoacusticus, hjernenerve nr. 8, er ligevægts- og hørenerven. | |
| N. olfactorius har trofisk centrum i bipolare nerveceller (4), der ligger i epitelet i næseslimhinden, nærmere betegnet den del, vi kalder for lugteområdet, regio olfactoria (1). Hele nerven består af ca 20 “lugtetråde”, fila olfactoria. De består af bundter af aksoner (2), der har apparent udspring fra lugtekolben (3), bulbus olfactorius. Den ligger på underfladen af hjernens lobus frontalis hen over sipladen, lamina cribrosa (5). Fila olfactoria perforerer dura og går gennem lamina cribrosa til næsehulen. | |
| Nervetrådene (2) i fila olfactoria danner som sagt synapse med celler i bulbus olfactorius, hvorfra neuriterne fortsætter gennem tractus olfactorius til centrale dele af rhinencephalon (6). Det corticale center for lugt ligger på medialfladen af lobus temporalis (4). Fila olfactoria er enestående på flere måder. For det første derved at nervecellerne ligger i en slimhindes epitelbeklædning. For det andet fornys cellerne hele tiden i modsætning til nerveceller alle andre steder. | |
| N. opticus er ikke en egentlig nerve, men en hjerneudløber. Det viser sig bl a ved, at nervetrådene har myelinskede, men mangler Schwann’s skede og derfor ikke kan regenerere. N. opticus ligner dermed hvid substans i hjernen. Trådene i n. opticus har trofisk centrum i multipolare nerveceller (1) i nethinden. Nerven forlader øjeæblet ved bageste pol og går igennem foramen opticum (2) til cavitas cranii. | |
| N. opticus (2) er omgivet af alle tre hjernehinder (3). Det betyder, at spalterne mellem hjernehinderne, fx subarachnoidalrummet mellem pia mater og arachnoidea, omgiver n. opticus helt frem til området, hvor nerven passerer gennem øjeæblets senehinde, sclera. | |
| Højre og venstre synsnerve (2) løber sammen i chiasma opticum (3), hvor de mediale tråde (4) fra højre og venstre side krydser hinanden |
Side 355
| i midtlinien, mens de laterale tråde (5) i de to nerver fortsætter uden at krydse. Krydsede og ukrydsede tråde danner tractus opticus (6). | |
| Tractus opticus (6) fortsætter til corpus geniculatum laterale (7). Her danner langt de fleste tråde synapse med multipolare nerveceller. Neuriterne fra de nye celler danner et samlet bundt (8), der går til synscentret, area 17 på lobus occipitalis (9). Andre tråde går fra corpus geniculatum laterale til øvre firhøje i mesencephalon (10), hvorfra der kan dannes refleksbuer til kerner for bl a øjeæblets bevægenerver. | |
| N. vestibulocochlearis er rent sensitiv. Den består af ligevægtsnerven, n. vestibularis (1), der har trofisk centrum i et ganglie (3) i bunden af meatus acusticus internus, og hørenerven, n. cochlearis (2), der har trofisk centrum i et ganglie i sneglen (6). Begge ganglier består af bipolare nerveceller. | |
| N. vestibulocochlearis har apparent udspring i furen mellem pons og medulla oblongata. Den løber til det indre øre gennem porus acusticus internus (4) og meatus acusticus internus, hvor dura perforeres. N. vestibularis fører impulser fra sanseepitelet i buegange (5) og forgårdssække (7), og n. cochlearis fra sanseepitelet i sneglen (6). Nervetrådene går til kerner (8) i hjernestammen. | |
| De tre “sansenerver”, hjernenerve nr. 1, 2 og 8, er forskellige fra de øvrige hjernenerver. De er desuden indbyrdes forskellige. N. olfactorius har trofisk centrum i nerveceller, der ligger i slimhindens epitel i næsehulen. Lugtecellerne er desuden de mest eksponerede, dvs overfladisk beliggende af kroppens nerveceller, og kan nydannes ved celledeling fra stamceller efter fødselen. | |
| N. opticus har trofisk centrum i nerveceller i nethinden, og dens tråde svarer til nervetråde i hvid substans i hjerne og rygmarv. Særlig vigtigt er det, at nervetrådene i n. opticus ikke kan regenerere, da Schwann’s celler mangler. Under forløbet i orbita fra foramen opticum til øjeæblets bagflade er n. opticus omgivet af alle tre hjernehinder med tilhørende spalteformede hulrum. |
Side 356
| N. vestibulocochlearis minder mest om en “almindelig” sensitiv nerve. Der er dog den mindre forskel, at nervetrådene har trofisk centrum i bipolare ganglieceller og ikke pseudounipolare celler, som ellers er det normale for sensitive ganglier. |
| De tre nerver er motoriske og omtales under ét. | |
| De har trofisk centrum i hver sin kerne (3), (4), (6) i hjernestammen og innerverer muskler, der bevæger øjeæblet. N. oculomotorius fører endvidere autonome, parasympatiske tråde, der har trofisk centrum i en parasympatisk kerne (3) (skraveret) i hjernestammen. | |
| N. oculomotorius (1) har apparent udspring på forsiden af mesencephalon i fordybningen mellem hjernestilkene, crura cerebri. N. trochlearis (2) har apparent udspring lige under de nedre firhøje, og det vil sige på bagsiden af hjernestammen, i modsætning til alle andre hjernenerver. N. abducens (3) har apparent udspring i furen mellem pons og medulla oblongata. | |
| N. oculomotorius, n. trochlearis og n. abducens løber fremad i kraniebunden, perforerer dura mater og fortsætter gennem fissura orbitalis superior (4) til orbita. | |
| N. trochlearis (2) innerverer den øverste skrå øjenmuskel (5), m. obliquus superior. Musklen skifter retning under sit forløb. Det sker ved hjælp af en senering, en trochlea, nær loftet ved den mediale øjenkrog. Heraf navnet, n. trochlearis. | |
| N. abducens (3) innerverer den laterale øjenmuskel (6), m. rectus lateralis, der drejer øjeæblet udad, “abducerer”. | |
| N. oculomotorius (1) innerverer de resterende ydre øjenmuskler. Den har endvidere parasympatiske tråde til ringmusklen, der forsnævrer pupillen (7), m. sphincter pupillae, og til akkomodationsmusklen (8), m. ciliaris. De parasympatiske impulser går gennem det lille parasympatiske ggl. ciliare (9), der ligger bag øjeæblet. |
Side 357
| Ved læsion af en af nerverne til øjenmusklerne optræder dobbeltsyn, fordi øjeæblet i den lammede side ikke kan følge det “raske” øjeæbles bevægelser. Når man læser, kan man fremkalde dobbeltsyn hos sig selv ved at trykke let mod det ene øjeæble, så man nedsætter bevægeligheden. |
| N. trigeminus, ansigtets store følenerve, er hjernenerve nr. 5. Den fører sensitive og motoriske tråde. De (fleste) sensitive tråde har trofisk centrum i ggl. trigeminale (9), kroppens største sensitive ganglie. Det indeholder pseudounipolare nerveceller ligesom hovedparten af sensitive ganglier. | |
| Neuriterne fra ggl. trigeminale fortsætter centralt til de sensitive trigeminuskerner (sorte) (5), der strækker sig gennem hele hjernestammen. Motoriske nervetråde i n. trigeminus har alle trofisk centrum i den motoriske trigeminuskerne i hjernestammen (hvid) (5). | |
| Det apparente udspring for n. trigeminus (V) ligger på forfladen af pons. Nervetrådene i n. trigeminus kommer frem i to bundter, et stort radix sensoria, der består af sensitive tråde, og et lille radix motoria (ikke tegnet), der består af motoriske tråde. | |
| Radix sensoria og radix motoria løber fremad og henover øverste og bageste kant af pars petrosa nær spidsen. Her ligger de i impressio trigeminalis omgivet af dura mater, og det er her, at radix sensoria (1) svulmer op og danner det største sensitive hjernenerveganglie, ggl. trigeminale (9). Fra gangliet afgår tre grene (3), (4) og (5). | |
| Mest medialt ligger første trigeminusgren (3), n. ophthalmicus. Den midterste er anden trigeminusgren (4), n. maxillaris. Længst lateralt ligger tredie trigeminusgren, n. mandibularis (5), der dannes af en gren fra radix sensoria (1) og hele radix motoria (2). | |
Side 358
| De sensitive tråde i n. trigeminus innerverer ansigtshud inden for områderne (6), (7) og (8), dvs fra det øverste punkt på hovedet, vertex (stort pilehoved), til basis mandibulae, orbita, cavitas nasi og cavitas oris. | |
| Grænsen mellem innervationsområdet for første (6) og anden (7) trigeminusgren går gennem øjenspalten, og grænsen mellem anden (7) og tredie (8) trigeminusgren går gennem mundspalten. Områderne (6), (7) og (8) kaldes tilsammen “trigeminusmasken”. Læg især mærke til, at maskens kant løber langs forkanten af m. masseter (pilehoveder ved 8), at den deler ydre øre i en øvre og nedre del og fortsætter til vertex, det højeste punkt på hovedet (stort pilehoved). | |
| Ud fra innervationen af hudområderne kan man gætte sig til de dybere liggende innervationsområder. I orbita innerveres indholdet af første trigeminusgren (6). Næsehulen “deles” mellem første (6) og anden (7) trigeminusgren, og mundhulen “deles” mellem anden (7) og tredie (8) trigeminusgren. | |
| De motoriske tråde i n. trigeminus innerverer de fire tyggemuskler, to muskler i mundbunden, m. mylohyoideus og venter anterior m. digastrici, én ganemuskel, m. tensor veli palatini, og én muskel i trommehulen, m. tensor tympani. | |
| Ggl. trigeminale (9) er et stort fladt ganglie. Det ligger i impressio trigemininalis (3) på øvre flade af pars petrosa. | |
| Gangliet ligger omgivet af en pose, der er dannet af dura. Posens åbning vender bagud mod fossa cranii posterior, og gennem åbningen løber radix sensoria og radix motoria ind i duraposen. Posens bund vender fremad, og dura følger de tre trigeminusgrene, til de forlader basis cranii interna gennem deres respektive huller. |
Side 359
| N. ophthalmicus (3) er første trigeminusgren. Den er rent sensitiv og afgår fra ggl. trigeminale (9). Den er rettet fremad og går ind i øjenhulen gennem fissura orbitalis superior. I loftet af orbita deler den sig straks i tre grene. Længst lateralt ligger n. lacrimalis (10). Den følger lateralvæggen i orbita. N. frontalis (11) løber midt i loftet af orbita, og n. nasociliaris (12) følger medialvæggen. Alle tre grene passerer gennem aditus orbitalis og ender med at forsyne huden (6). | |
| N. lacrimalis (10) forsyner først og fremmest tårekirtlen, gl. lacrimalis. N. supraorbitalis (11a) er den længste gren fra n. frontalis. Den løber op over panden og fortsætter helt til vertex (pilehoved). N. nasociliaris (12) ender med grene til forreste del af næsehulen og huden på ydre næse. | |
| N. ophthalmicus innerverer dermed øjeæble og tårekirtel (sensitive tråde), øjelåg (øvre) (6), huden på næsen og i panden (6) (til vertex) og slimhinden i forreste del af næsehulen. | |
| Til n. ophthalmicus er knyttet det lille parasympatiske ggl. ciliare | |
| Det ligger bag øjeæblet, og gennem gangliet (9) løber parasympatiske impulser fra n. oculomotorius (1) til ringmusklen, der forsnævrer pupillen (7), m. sphincter pupillae, og til akkomodationsmusklen (8), m. ciliaris. Ggl. ciliare (9) er omkoblingsstation for parasympatiske tråde fra n. oculomotorius (1). | |
| Ganglion ciliare har intet med tårekirtlens sekretion at gøre. Gl. lacrimalis (9) modtager parasympatiske, sekretoriske, nervetråde fra n. facialis (2), og de er omkoblet i et andet parasympatisk ganglie (8), ggl. pterygopalatinum, og ikke i ggl. ciliare (5). Det er n. lacrimalis, der indeholder trådene det sidste stykke ud til tårekirtlen. |
| N. maxillaris er anden trigeminusgren. Den er rent sensitiv og afgår fra ggl. trigeminale (9). N. maxillaris forlader cavitas cranii gennem foramen rotundum (1). Den fortsætter gennem fossa pterygopalatina (2) og videre fremad gennem fissura orbitalis inferior (3) til orbita. Her løber den i bun- |
Side 360
| den, først i sulcus infraorbitalis og derefter i canalis infraorbitalis, der åbner sig på forfladen af maxilla med foramen infraorbitale (4). N. maxillaris skifter navn til n. infraorbitalis, idet den passerer gennem fissura orbitalis inferior (3). | |
| N. maxillaris og n. infraorbitalis innerverer overkæben med tænder og gingiva, hud og slimhinde ud for overkæbe og kindben (7), dvs hud og slimhinde på overlæbe, huden på en del af næse, kind og nedre øjelåg, endvidere slimhinden på ganen og i det meste af næsehulen, huden i forreste fjerdedel af regio temporalis (pil). | |
| Grene fra n. maxillaris afgår i fossa pterygopalatina, som de forlader gennem forskellige huller. Grenene er rami nasales (5), nn. palatini (6) og rami alveolares superiores posteriores (7c). | |
| Rami nasales (1) består af nervetråde fra n. maxillaris (3). Trådene (2) løber fra n. maxillaris gennem det parasympatiske ganglie, ggl. pterygopalatinum (4), og fortsætter gennem foramen sphenopalatinum (ved 1) til næsehulen. | |
| Så snart de er kommet gennem foramen sphenopalatinum (1) og ind i næsehulen, deler de sig i laterale grene (2) til bageste del af lateralvæggen (3), og i mediale grene til skillevæggen (2). | |
| Den længste gren her hedder n. nasopalatinus (3), og den løber fra næseskillevæggen gennem canalis incisivus (4) og foramen incisivum til allerforreste del af den hårde gane, som den innerverer sammen med den palatinale gingiva ud for de fire fortænder. N. nasopalatinus skal bedøves, før en fortand i overkæben trækkes ud. | |
| Nn. palatini (5) består også af nervetråde fra n. maxillaris (3). Trådene (2) løber gennem det parasympatiske ggl. pterygopalatinum (4) og videre mod nederste spids af fossa pterygopalatina. | |
| Nn. palatini (3) og (4) forlader spidsen af fossa pterygopalatina gennem kanaler, der åbner sig på ganen gennem foramina palatina. | |
| N. palatinus major (4) går gennem foramen palatinum majus (5) og løber fremad på den hårde gane til linien mellem hjørnetændernes mesialflader, dvs de to flader, der vender fremad mod fortænderene. Her danner den anastomose (3) med n. nasopalatinus (2), der løber gennem foramen incisivum (1). |
Side 361
| N. palatinus major innerverer den hårde gane og palatinale gingiva bag hjørnetændernes mesialflader. Nn. palatini minores (6) løber gennem foramina palatina minora til den bløde gane. | |
| Rami alveolares superiores posteriores (2) afgår fra n. maxillaris (1) i fossa pterygopalatina, som de forlader gennem den laterale, trekantede åbning (3). De fortsætter på facies infratemporalis maxillae til foramina alveolaria (4). Før de passerer gennem hullerne, afsender de rami gingivales (5), der fortsætter til gingiva vestibulært (facialt) for molarerne i overkæben. | |
| Når rami alveolares superiores posteriores er gået igennem foramina alveolaria, ligger de i bagvæggen af kæbehulen mellem knogle og slimhinde. De (7c) går til plexus dentalis superior (8) over tænderne og innerverer herigennem overkæbens molarer. Rami alveolares superiores posteriores bedøves gennem mundhulen, ved at føre en kanyle op langs facies infratemporalis maxillae og sprøjte bedøvelsesvæsken ud omkring nerverne her. | |
| Grene fra n. infraorbitalis er ramus alveolaris superior medius (7b) og rami alveolares superiores anteriores (7a). Af endegrene nævner vi kun rami labiales superiores (11). Ramus alveolaris superior medius (7b) afgår fra n. infraorbitalis i sulcus infraorbitalis, mens rami alveolares superiores anteriores (7a) afgår en lille centimeter inde i canalis infraorbitalis, regnet fra foramen infraorbitale (4). Alle rami alveolares går til plexus dentalis superior (8). Rami alveolares superiores anteriores (7a) kan bedøves, ved at føre en kanyle gennem foramen infraorbitale (4) og sprøjte bedøvelsesvæske omkring n. infraorbitalis i kanalen. Ramus alveolaris superior medius kan ikke nås gennem kanalen. I stedet lægges bedøvelsen i slimhinden ud for rodspidsen af præmolarerne. | |
| N. infraorbitalis (5) forlader canalis infraorbitalis gennem foramen infraorbitale (6) og deler sig straks i sine endegrene, hvor de vigtigste er rami labiales superiores (1). | |
| De innerverer overlæbens hud og slimhinde (1), slimhinde i vestibulum oris og gingiva vestibulært, dvs facialt, for alle tænderne i overkæben bortset fra molarerne, hvor gingiva innerveres af rami gingivales fra rami alveolares superiores posteriores. | |
Side 362
| Til n. maxillaris er knyttet det parasympatiske ggl. pterygopalatinum (10). | |
| Gangliet (4) ligger under nerven (3) i fossa pterygopalatina, og sensitive tråde (2) fra n. maxillaris går gennem gangliet og til rami nasales (1) og nn. palatini (5). | |
| N. mandibularis er tredie trigeminusgren. Den indeholder både sensitive og motoriske tråde. De sensitive tråde kommer fra ggl. trigeminale, og de motoriske fra radix motoria. | |
| N. mandibularis går igennem foramen ovale (1), og den kommer ud i spatium lateropharyngeum (3) umiddelbart bag pterygoidermuren (5) og deler sig straks i en masse grene. | |
| Set lateralt fra skjuler pterygoidermuren (fig. 16) (5) delingsstedet af n. mandibularis i spatium lateropharyngeum. Fjerner vi den øverste del af pterygoidermuren (fig. 17), dvs m. pterygoideus lateralis, kan vi se nervens delingssted lige under foramen ovale i spatium lateropharyngeum, bagved “muren”. | |
| Læg mærke til, at den bageste grænse for “trigeminusmasken”, og dermed for n. mandibularis, følger forkanten af m. massseter (pilehoveder lige bag 8) og går henover øreflippen. De motoriske tråde i n. mandibularis innerverer alle fire tyggemuskler, to muskler i mundbunden, m. mylohyoideus og venter anterior af m. digastricus |
Side 363
| Grenene fra n. mandibularis begynder alle i spatium lateropharyngeum. De fleste af grene går herfra igennem pterygoidermurens spalter og ind i fossa infratemporalis. Hovedparten bruger fossa infratemporalis som “gennemgangsregion” og forlader den igen ad forskellige veje. Grenene til mm. pterygoidei bliver dog i fossa infratemporalis, hvor musklerne ligger. | |
| To små grene (20) og (21) fra n. mandibularis bliver i spatium lateropharyngeum, hvor alle nervens grene begynder. Den ene (20) er grenen til den ganemuskel, der innerveres af n. mandibularis, m. tensor veli palatini. Den anden (21) er grenen til en muskel, der ender i trommehulen, m. tensor tympani. | |
| Endelig løber en enkelt gren (14) lateralt, idet den følger pterygoidermurens bagside til regio parotidea, og derfra til regio temporalis. |
| N. mandibularis afgiver fire store sensitive grene n. alveolaris inferior, “underkæbenerven” (2), n. lingualis, “tungenerven” (8), n. buccalis, “kindnerven” (12) og n. auriculotemporalis, “øretindingenerven” (14). | |
| N. alveolaris inferior, “underkæbenerven” (2), går fra spatium lateropharyngeum til fossa infratemporalis. Efter et kort forløb forlader den fossa infratemporalis gennem foramen mandibulae og fortsætter i canalis mandibulae (6). Her sender den grene (7) til alle tænder i underkæben. | |
| Under det fremadrettede forløb i canalis mandibulae afsender n. alveolaris inferior desuden n. mentalis (8), der løber gennem foramen mentale. Nerven innerverer hagens hud (11) og underlæbens hud og slimhinde (12). Desuden slimhinden i vestibulum oris med gingiva ud for fortænder, hjørnetand og præmolarer. | |
| N. lingualis, “tungenerven” (8), går fra spatium lateropharyngeum gennem pterygoidermuren til fossa infratemporalis. Nerven (8) |
Side 364
| forlader fossa infratemporalis (4) ved forkanten af m. pterygoideus medialis (pilehoveder) og fortsætter nedad og medialt for corpus mandibulae til trigonum submandibulare (28). Her har nerven et kort fremadrettet forløb, hvorefter den løber henover m. mylohyoideus (11) og ind i regio sublingualis (10). N. lingualis innerverer slimhinden på forreste del af tungen og i mundbunden samt linguale gingiva i underkæben. | |
| N. buccalis, “kindnerven” (12), går fra spatium lateropharyngeum gennem pterygoidermuren til fossa infratemporalis. Den forlader fossa infratemporalis ved at følge lateralfladen af m. buccinator (13). På den måde kommer n. buccalis ind i regio buccalis, hvor den innerverer hud og slimhinde samt faciale gingiva ud for molarerne i underkæben. | |
| N. auriculotemporalis, “øretindingenerven” (14), afgår fra n. mandibularis i spatium lateropharyngeum. Den går ikke gennem pterygoidermuren, men følger i stedet bagfladen af musklerne, her repræsenteret af m. pterygoideus lateralis, lateralt til gl. parotidea. | |
| N. mandibularis afgiver en række motoriske grene, nerven til m. masseter (15), nerverne til m. temporalis (17), nerverne til mm. pte |
Side 365
| rygoidei (19) og (19a), nerven til m. tensor veli palatini (20) og nerven til m. tensor tympani (21). De motoriske nervetråde til m. mylohyoideus og venter anterior m. digastrici skal kendes, men deres forløb gennemgås ikke i basismodulet. | |
| Nerven til m. masseter (15) begynder i spatium lateropharyngeum, hvor den afgår fra n. mandibularis. Nerven løber fremad til fossa infratemporalis gennem pterygoidermuren (5) øverst, under basis cranii. Den forlader fossa infratemporalis ved at gå gennem incisura mandibulae (16) til regio masseterica, hvor den innerverer m. masseter. | |
| Nerverne til m. temporalis (17) afgår fra n. mandibularis i spatium lateropharyngeum. De løber fremad gennem pterygoidermuren ind i fossa infratemporalis, hvor forløbet er kort, inden de forlader regionen gennem den store ubenævnte åbning medialt for arcus zygomaticus og går til regio temporalis. Her innerverer de m. temporalis. | |
| Nerverne til mm. pterygoidei (19) har et ganske kort forløb fra n. mandibularis i spatium lateropharyngeum til mm. pterygoidei i fossa infratemporalis. Det er de eneste grene fra n. mandibularis, der ender i fossa infratemporalis, hvor mm. pterygoidei ligger. Nerverne til mm. pterygoidei nævnes under ét, selv om der selvfølgelig er nerver til henholdsvis m. pterygoideus lateralis og medialis. | |
| Nerven til m. tensor veli palatini (20) afgår fra n. mandibularis i spatium lateropharyngeum, og den bliver i spatium lateropharyngeum, idet m. tensor veli palatini udspringer her. Nerven er tynd og kort. Nerven til musklen i mellemøret m. tensor tympani (21), afgår fra n. mandibularis i spatium lateropharyngeum. Den bliver i spatium lateropharyngeum, idet den har et kort forløb til canalis musculotubarius, hvor musklen udspringer. | |
| Til n. mandibularis er knyttet to parasympatiske ganglier, ggl. oticum (22) og ggl. submandibulare (23). | |
| Ggl. oticum ligger tæt på bagsiden af n. mandibularis i spatium lateropharyngeum. Gangliet ligger lige under foramen ovale (1). I ggl. oticum danner parasympatiske, sekretoriske nervetråde fra hjernenerve nr. 9, n. glossopharyngeus, synapse med små multipolare nerveceller, hvorfra andet neuron fortsætter til gl. parotidea (P). |
Side 366
| Neuriterne i andet neuron følges med nervetrådene i n. auriculotemporalis, der netop løber fra n. mandibularis til regio parotidea på sin vej til regio temporalis. | |
| Ggl. submandibulare (23) ligger ophængt under n. lingualis (8) i trigonum submandibulare (28). Gennem gangliet løber parasympatiske, sekretoriske impulser fra hjernenerve nr. 7, n. facialis til gl. submandibularis og gl. sublingualis. | |
| N. facialis, ansigtets bevægenerve, er hjernenerve nr. 7. Den fører motoriske, parasympatiske og sensitive tråde, bl a smagstråde. | |
| De motoriske tråde har trofisk centrum i den motoriske facialiskerne (7) (hvid) i hjernestammen, og de parasympatiske tråde har trofisk centrum i den parasympatiske facialiskerne (7) (skraveret), der også ligger i hjernestammen. De sensitive tråde i n. facialis har trofisk centrum i et lille sensitivt ganglie, ggl. geniculi, knyttet til n. facialis, mens nerven løber gennem pars petrosa. Neuriterne fra gangliets pseudounipolare nerveceller ender i en sensitiv kerne (7, 9, 10) (sort) i hjernestammen. | |
| Kernen (5) er fælles for smagstrådene i n. facialis (3), n. glossopharyngeus (2) og n. vagus (1) og hedder nucleus tractus solitarii. | |
| Det apparente udspring for n. facialis (VII) er i furen mellem pons og medulla oblongata. | |
| Fra udspringsstedet (1) løber nerven frem gennem porus og meatus acusticus internus (2) sammen med n. vestibulocochlearis. N. facialis perforerer dura i bunden af meatus acusticus internus. |
Side 367
| Fra meatus acusticus internus løber n. facialis videre gennem sin egen kanal (3) i pars petrosa. | |
| Kanalen hedder canalis facialis og ender ved foramen stylomastoideum (4) på nedre flade af pars petrosa. | |
| Da foramen stylomastoideum ligger i loftet af regio parotidea (4), betyder det, at n. facialis kommer ud i denne region, der også kaldes regio retromandibularis. I kirtlen danner nerven et netværk, og herfra afgår dens fem sæt endegrene. Grenene forlader gl. parotidea og fortsætter i subcutis til ansigtsmusklerne. | |
| Nogle af endegrenene kaldes rami buccales (4). De ligger først på lateralfladen af m. masseter, i subcutis, uden på fascien. Denne overfladiske lejring i regio masseterica medfører, at dybe snitsår her kan lædere facialisgrene, ligesom kulde og træk kan påvirke grenene. I gl. parotidea ligger grenene mere beskyttet. | |
| N. facialis innerverer alle mimiske muskler, dvs ansigtsmuskler, platysma (5), venter posterior m. digastrici (6), | |
| m. stylohyoideus (6) og m. stapedius (4), stigbøjlens muskel i mellemøret. | |
| I n. facialis løber parasympatiske, sekretoriske tråde. Trådene fører impulser til kirtler i tre store hulheder, orbita, cavitas nasi og cavitas oris samt til to store spytkirtler. Det drejer sig om |
Side 368
| gl. lacrimalis i orbita (7), gll. nasales i cavitas nasi (8), gll. palatinae i cavitas oris (9), gl. submandibularis (10) og gl. sublingualis (11). Det vil sige, at gl. parotidea er den eneste store spytkirtel, der ikke får sekretoriske impulser fra n. facialis. Den får fra n. glossopharyngeus. | |
| De parasympatiske impulser til tårekirtel, næsekirtler og ganekirtler går gennem ggl. pterygopalatinum (12). Impulserne til gl. submandibularis og gl. sublingualis går gennem ggl. submandibulare (13). | |
| De parasympatiske impulsers vej til kirtlerne i orbita, næsehule og gane begynder i den parasympatiske facialiskerne (2), og neuriterne fra kernens celler går til ggl. pterygopalatinum (8). Her danner de synapse med små multipolare nerveceller, og neuriterne fra disse celler, dvs andet neuron, går til gl. lacrimalis (9), gll. nasales (10) og gll. palatinae (11), idet trådene følges med nervegrene fra n. maxillaris og n. ophthalmicus. | |
| Andre tråde fra den parasympatiske facialiskerne går via chorda tympani til ggl. submandibulare (12). Her danner de synapse, og andet neuron fortsætter til gl. submandibularis (14) eller gl. sublingualis (13). | |
| N. facialis (3) indeholder smagstråde fra pars presulcalis af dorsum linguae. Det er papillae fungiformes, der bærer smagsløgene. De store papillae vallatae lige foran sulcus terminalis innerveres fra n. glossopharyngeus. Smagstrådene fra papillae fungiformes følger først n. lingualis og løber derefter via chorda tympani til n. facialis. Trofisk centrum for første neuron ligger i ggl. geniculi (4), og neuriten fra den pseudounipolare nervecelle ender i hjernestammen i nucleus tractus solitarii (5), hvor den danner synapse med 2. neuron. |
Side 369
| N. glossopharyngeus er hjernenerve nr. 9. Den fører sensitive, parasympatiske og motoriske tråde. | |
| De sensitive tråde i n. glossopharyngeus har trofisk centrum i små ganglier under basis cranii. Gangliecellerne er pseudounipolare, og deres neuriter går til en sensitiv kerne i hjernestammen (7), (9), (10) (sort). Kernen er fælles for n. facialis, n. glossopharyngeus og n. vagus og hedder nucleus tractus solitarii. De parasympatiske tråde i n. glossopharyngeus har trofisk centrum i en lille parasympatisk kerne (9) (skraveret) i hjernestammen. | |
| De motoriske tråde i n. glossopharyngeus har trofisk centrum i en motorisk kerne i hjernestammen (9, 10, 11) (hvid). Kernen er fælles for n. glossopharyngeus, n. vagus og n. accessorius. | |
| Det apparente udspring for n. glossopharyngeus (IX) er på forfladen af medulla oblongata. Her kommer både de sensitive, parasympatiske og motoriske tråde til syne. | |
| N. glossopharyngeus perforerer dura mater lige før, den forlader kraniehulen gennem foramen jugulare (1). Nerven kommer ud i spatium lateropharyngeum (2), som den gennemkrydser. | |
| lateropharyngeum løber n. glossopharyngeus mellem de to store arterier, a. carotis interna og externa (4) og (3). Dens ledestruktur her er m. stylopharyngeus (6). | |
| N. glossopharyngeus forlader spatium lateropharyngeum ved at perforere svælgvæggen (ved stiplingen). Den ender bagtil i tungen (7). | |
| De sensitive tråde i n. glossopharyngeus innerverer slimhinden på bageste del af tungen (7) og svælg (8) (med tonsiller). Desuden arterievæg ved delingsstedet af den store halsarterie, a. carotis communis (5). De sensitive tråde til arterievæggen registrerer ændringer i blodtryk og blodets iltindhold. N. glossopharyngeus er den egentlige smagsnerve. Den fører smagstråde (2) fra papillae foliatae og papillae vallatae, og da hver eneste papilla vallata har flere hundrede smagsløg, betyder det, at flertallet af smagsløgene har deres nerveforsyning gennem n. glossopharyngeus. |
Side 370
| De parasympatiske impulser fra n. glossopharyngeus løber til det parasympatiske ggl. oticum (11), hvor der er synapse. Andet neuron fortsætter til gl. parotidea (10). | |
| Ggl. oticum (22) ligger under foramen ovale (1), tæt på bagfladen af n. mandibularis, i spatium lateropharyngeum. | |
| Fra gangliet er det naturligt, at de parasympatiske tråde fortsætter i n. auriculotemporalis, der netop løber lateralt til gl. parotidea, inden den fortsætter opad til regio temporalis. De motoriske tråde i n. glossopharyngeus innerverer nervens ledemuskel, m. stylopharyngeus (6). | |
| N. vagus, hjernenerve nr. 10, er den hjernenerve, der når længst væk fra sit udspring. | |
| N. vagus fører sensitive, parasympatiske og motoriske tråde. Det er den nerve, der fører det største antal parasympatiske tråde. Den indeholder flere parasympatiske tråde end de andre nerver tilsammen. I absolutte tal er der dog flest sensitive tråde. | |
| De parasympatiske tråde i n. vagus har trofisk centrum i den parasympatiske vaguskerne (10) (skraveret), og de motoriske tråde har trofisk centrum i en motorisk kerne (9, 10, 11) (hvid). Den motoriske kerne er fælles for n. glossopharyngeus, n. vagus og n. accessorius, men de fleste celler i kernen hører til n. vagus. Den parasympatiske og motoriske kerne ligger begge i hjernestammen. |
Side 371
| De sensitive vagustråde har trofisk centrum i sensitive vagusganglier lige under foramen jugulare i basis cranii. Neuriterne fra de psudounipolare nerveceller går til en sensitiv kerne i hjernestammen (7, 9, 10) (sort). Kernen er fælles for n. facialis, n. glossopharyngeus og n. vagus og hedder nucleus tractus solitarii. | |
| N. vagus (X) har sit apparente udspring på forfladen af medulla oblongata. | |
| N. vagus perforerer dura mater, før den forlader kraniehulen gennem foramen jugulare (1) og kommer ud i spatium lateropharyngeum. Den løber lodret ned gennem hele spatium lateropharyngeum (2) fra basis cranii externa til apertura thoracis superior. Derefter fortsætter n. vagus videre gennem hele cavitas thoracis. Først ligger den i mediastinum superius (3) langs luftrøret. Derefter i mediastinum posterius (4), hvor det er oesophagus (6), som er nervens “ledestruktur”. N. vagus løber bagved lungeroden (12) og perforerer diafragma sammen med oesophagus ved (5). Den forgrener sig til slut i bughulen under diafragma (7). | |
| De parasympatiske tråde i n. vagus innerverer viscera i hals, brysthule og størstedelen af bughulen. De parasympatiske tråde i n. vagus spiller en enorm rolle. Det er fx impulser gennem disse tråde, der sørger for, at hverken hjerte eller respiration går for hurtigt. Det er også impulser fra den parasympatiske vaguskerne, der sætter gang i peristaltik i tarmen, og som er med til at fremme sekretion i ventrikel og pancreas. Ejendommeligt nok virker n. vagus som “bremse” på funktionerne i brysthulen og som “igangsætter” af funktioner i bughulen. Det sympatiske nervesystem har den modsatte virkning. |
Side 372
| De motoriske tråde i n. vagus innerverer tværstribet indvoldsmuskulatur. Det drejer sig om ganens muskler (8), undtagen m. tensor veli palatini, svælgets muskler (9), undtagen m. stylopharyngeus, larynxmuskler (10) (samtlige) og tværstribet muskulatur i oesophagus (6). | |
| De sensitive tråde i n. vagus forsyner slimhinder i fordøjelseskanalen, fra bageste del af tungen til flexura coli sinistra midt på tyktarmen og i luftvejene fra aditus laryngis til lungealveolerne. Til de sensitive tråde hører også enkelte smagstråde (1) fra området omkring vallecula epiglottica. | |
| Da n. vagus innerverer den tværstribede muskulatur i gane, svælg og strubehoved, betyder det, at en vaguslæsion kan medføre lammelse af “ganesejl” og svælg med synkebesvær til følge, samt lammelse af strubehovedet med respirations- og talebesvær til følge. | |
| N. accessorius, hjernenerve nr. 11, er “binerve” for n. vagus. Den er motorisk. Nogle tråde har trofisk centrum i den motoriske kerne i hjernestammen (9, 10, 11), der er fælleskerne for n. glossopharyngeus, n. vagus og n. accessorius. Andre tråde i n. accessorius har trofisk centrum i forhornsceller i øverste del af medulla spinalis. | |
| N. accessorius består altså af en hjernenervedel med udspring i en hjernenervekerne og en rygmarvsnervedel med udspring i forhorn. Derfor har n. accessorius to apparente udspring. Det ene (1) på forfladen af medulla oblongata og det andet (2) på halsafsnittet af medulla spinalis. Trådene (2) løber først op gennem foramen magnum |
Side 373
| til cavitas cranii, hvorefter de følges med trådene (1) til foramen jugulare (3). Her perforerer nerven dura mater, lige før den går igennem hullet til spatium lateropharyngeum (5). | |
| Trådene (1) i n. accessorius fra medulla oblongata “går over på” n. vagus (4) og fortsætter sammen med denne. Trådene (2) fra medulla spinalis går selvstændigt til m. sternocleidomastoideus (6) og m. trapezius (7). | |
| N. hypoglossus (1), tungens bevægenerve, er hjernenerve nr. 12. | |
| Den er motorisk, og trådene har trofisk centrum i hypoglossuskernen (12) i hjernestammen. | |
| Nervens apparente udspring (XII) er på forfladen af medulla oblongata. Den perforerer dura mater og går straks derefter gennem canalis hypoglossi (10). | |
| Kanalen åbner sig i spatium lateropharyngeum (2), og her løber n. hypoglossus (1) i en bue nedad, fremad og lateralt for begge de store arterier (3) og (4), a. carotis externa og interna. | |
| Kort efter at n. hypoglossus (1) har krydset “carotiderne” (3) og (4), forlader den spatium lateropharyngeum (2). Det sker, når nerven passerer profundt for mellemsenen af m. digastricus (5) og fortsætter ind i trigonum submandibulare, hvor den (6) har et kort forløb. Fra trigonum submandibulare løber n. hypoglossus fremad og ind over m. mylohyoideus (7) til regio sublingualis (8). Her fortsætter den op i tungen (9). | |
| N. hypoglossus innerverer tungemuskler undtagen to, musklen mellem tunge og gane, m. palatoglossus, og musklen mellem tunge og svælg, pars glossopharyngea af m. constrictor pharyngis superior. |
Side 374
| Der er 31 par rygmarvsnerver, spinalnerver, nn. spinales. De har apparent udspring på sidefladen af medulla spinalis. Her udspringer hver rygmarvsnerve med to rødder, radix posterior (2), der er sensitiv, og radix anterior (1), der er motorisk. Radix posterior har lateralt en opsvulmning, ggl. spinale (3), der indeholder pseudounipolare nerveceller (1). | |
| Cellelegemerne er trofisk centrum for nervetrådene (3), dvs cellernes neuriter i radix posterior (5), og for nervetrådene (2), der går til periferien, dvs cellernes dendriter. | |
| De fleste tråde (6) i radix anterior har trofisk centrum i forhorn, dvs, det er motoriske tråde til tværstribede muskler. Et mindre antal tråde (7) har trofisk centrum i lateralhorn i rygmarvens brystdel, det er autonome, sympatiske tråde til glatte muskler og kirtler. | |
| De to radices, radix anterior og posterior, mødes lige lateralt for ggl. spinale og danner den korte spinalnerve (5). Nerven forlader canalis vertebralis gennem foramen intervertebrale (6), hvorefter den deler sig i to rami, ramus anterior (7) og ramus posterior (8). Begge rami er blandede, dvs, de indeholder både sensitive, motoriske og evt. autonome tråde. | |
| Rami anteriores er tykkere og længere end rami posteriores. Rami anteriores indeholder mange flere nervetråde end rami posteriores. Det er nødvendigt, da de forsyner et stort hudområde og mange muskler på kroppens forside samt ekstremiteterne. Rami posteriores forsyner kun ryggens hud og muskler. Undtagelsen fra denne regel er næstøverste spinal- eller cervicalnerve. | |
| Her er rami posteriores størst og kaldes n. occipitalis major, der innerverer nakken og bageste del af skalpen (1) til vertex. Husk at panden og det meste af ansigtet forsynes af n. trigeminus (hvide område). |
Side 375
| De 31 par spinalnerver omfatter (i højre og i venstre side): 8 cervicalnerver, 12 thoracalnerver, 5 lumbalnerver, 5 sacralnerver og 1 coccygealnerve. | |
| Inden for hver gruppe nummereres nerverne oppefra. | |
| Rami posteriores fra spinalnerverne løber bagud mellem rygmusklerne. De innerverer musklerne og fortsætter til huden på rygsiden. Den kraftigste ramus posterior i nakkeregionen er n. occipitalis major (3) fra cervicalnerve nr. 2. | |
| Den løber gennem nakkemusklerne til subcutis og fortsætter i dette lag til vertex, hvor den anastomoserer med n. supraorbitalis fra n. frontalis, der er en gren fra n. ophthalmicus (øverste hvide område). | |
| Rami anteriores af spinalnerverne anastomoserer og danner plexer (1) og (3). | |
| Det gælder dog ikke for thoracalnerverne, der fortsætter mellem ribbenene som enkeltnerver. | |
| Plexus cervicalis dannes af rami anteriores af de øverste cervicalnerver, og plexus brachialis dannes af rami anteriores af de nederste cervicervicalnerver. | |
| Plexus lumbalis (1) dannes af rami anteriores af lumbalnerverne, og plexus sacralis (3) dannes af rami anteriores af sacralnerverne. Plexus lumbalis (1) og plexus sacralis (3) hænger sammen. Undertiden beskrives de som et stort plexus lumbosacralis. |
Side 376
| Plexus cervicalis ligger i den øverste spids af scalenerporten (8), og i portens forlængelse opad. Scalenerporten (8) er trekantet med spidsen opad. I fortsættelse af spidsen er der en smal spalte mellem musklerne og det er her plexus cervicalis ligger. | |
| Plexus brachialis ligger i den nederste del af scalenerporten (1). Scalenerportens trekantede åbning begrænses af m. scalenus medius (6) bagtil, | |
| m. scalenus anterior (5) fortil og øverste ribben, costa 1, nedadtil. | |
| Grenene fra plexus cervicalis deles i superficielle og profunde. | |
| De superficielle grene fra plexus cervicalis kommer alle frem ved bagkanten af m. sternocleidomastoideus (1), midt på musklen. De superficielle grene (2‑5) innerverer huden i bageste del af regio temporalis, n. occipitalis minor (2), regio parotideomasseterica, n. auricularis magnus (3), regionerne på halsens forside (4) og nøglebens- og skulderregionerne (5). | |
| Hudområdet (2) på hoved og hals innerveres af de superficielle grene fra plexus cervicalis. | |
| Særlig interessant i tandlægepraksis er grænsen mellem hudområdet på hovedet og “trigeminusmasken” (6), (7) og (8). Læg især mærke til, at huden over m. masseter innerveres af n. auricularis magnus fra plexus cervicalis. Trigeminusmasken ender ved forkanten af m. masseter. | |
| De profunde grene af plexus cervicalis innerverer prævertebralmuskler, infrahyoide muskler (1), m. geniohyoideus (2) og diafragma (gennem 3). | |
| Nerven til diafragma er n. phrenicus (3). Den løber fra halsen gennem mediastinum superius (4) og derefter gennem mediastinum medium (5). Her krydser den foran lungeroden og løber langs hjerte (6) og hjertepose til mellemgulvet (7). |
| Plexus brachialis dannes af rami anteriores af de nederste cervicalnerver. Plexus brachialis går gennem scalenerporten (1) og fortsætter over første ribben (2) til armhulen (axillen) (3). Her afsender plexus brachialis de store grene til den frie del af overekstremiteten. | |
Side 377
| Rami anteriores fra thoracalnerverne løber som nn. intercostales mellem musklerne i ribbensmellemrummene. | |
| Plexus lumbalis (1) dannes under diafragma mellem musklerne på bageste bugvæg. De fleste grene løber nedad, og den tykkeste er n. femoralis (2) til lårets forside. N. femoralis løber sammen med arterien og venen af samme navn under lyskebåndet til lårets forside, hvor den innerverer hud og muskler. N. femoralis indeholder både sensitive (afferente) og motoriske (efferente) tråde, der indgår i knærefleksbuen. Er refleksen normal, vil knæet strækkes, når musklen kontraherer sig. Knærefleksen undersøges ved at slå på ligamentet mellem knæskal og skinneben. Derved strækkes muskeltenene i musklen på lårets forside, og sensitive (afferente) impulser fra tenene ledes til rygmarven, hvor de omkobles gennem synapser til motoriske neuroner. | |
| Plexus sacralis (3) ligger i bagvæggen af lille bækken på forfladen af os sacrum. Grenene fra plexus sacralis forlader bækkenhulen og løber gennem sæderegionen til lårets bagside. Den længste og tykkeste gren er n. ischiadicus (4). Den har grene til foden. | |
Side 378
| Den autonome del af nervesystemet omfatter sympaticus og parasympaticus. | |
| Det sympatiske nervesystem øger kroppens fysiske ydeevne og aktiveres ved forskellige former for stress, herunder fysisk krævende situationer. Det parasympatiske nervesystem har derimod størst aktivitet i normale hvilesituationer og stimulerer bl a fordøjelsen og opbygningen af kroppens reserver. Forskellen på sympaticus og parasympaticus udtrykkes præcist og enkelt på engelsk med ordene “fight and flight”, “rest and digest”. | |
| De fleste af kroppens organer er under autonom kontrol og modtager nervetråde fra såvel det parasympatiske som det sympatiske nervesystem. Der er nogle enkelte undtagelser fra dette mønster. De fleste blodkar modtager fx kun sympatiske nervetråde. I de tilfælde, hvor organerne har en dobbelt nerveforbindelse, er virkningen af de to dele af det autonome nervesystem som regel modsat. Øget aktivitet i de sympatiske nervetråde til hjertet medfører fx højere hjertefrekvens, mens øget aktivitet i de parasympatiske nervetråde nedsætter hjertefrekvensen. | |
| I mavetarmkanalen er rollerne byttet om. Impulser i de parasympatiske nervetråde medfører her øget aktivitet i de glatte muskelceller, der ytrer sig som øget peristaltik. Øget sympatisk aktivitet vil derimod få tarmmuskulaturen til at slappe af. | |
| De områder, som kun har forbindelse med det ene af de to autonome systemer, kan alligevel både stimuleres eller hæmmes. Det skyldes, at der selv i hvile løber en vis mængde af impulser i de autonome fibre, og denne aktivitet kan derfor både øges eller mindskes. Det er på den måde, at sympaticus regulerer diameteren på blodkarrene. Øges sympaticusaktiviteten, vil de glatte muskelceller trække karvæggen sammen og mindske diameteren, mens diameteren bliver større, når sympaticusaktiviteten bliver mindre. | |
| Sympaticus (7) udspringer fra lateralhorn i medulla spinalis. Lateralhorn findes kun i rygmarvens brystdel og to øverste lændesegmenter. | |
| Parasympaticus udspringer fra kerner i hjernestammen og fra rygmarvens sacraldel (1), (2), (3), (4) og (29). |
Side 379
| Hovedudspringsstedet for parasympaticus er kernerne i hjernestammen (1), (2), (3), (4). Ved at sammenligne tegningerne af det autonome nervesystem, kan man få det indtryk, at sympaticus og parasympaticus er lige store. Det er ikke tilfældet. Sympaticus er størst. |
| De sympatiske nervetråde går fra medulla spinalis til grænsestrengen, truncus sympaticus (1) i højre og venstre side. De sympatiske tråde forlader medulla spinalis gennem radix anterior sammen med de motoriske. Gennem anastomoser når de sympatiske tråde fra radix anterior til grænsestrengen i samme side. |
| Truncus sympaticus (1) er en nervestamme, der ligger langs hvirvelsøjlen (2), fra basis cranii externa til os coccygis (4). Der er kun ringe afstand mellem truncus sympaticus i højre og venstre side. Truncus sympaticus minder om en uregelmæssig “perlesnor”, fordi den hyppigt har opsvulmninger af ganglier, der varierer i størrelse. | |
| De fleste sympatiske tråde (7) fra lateralhornene danner synapse med små multipolare nerveceller (8) i grænsestrengsganglierne. Neuriterne fra de nye celler, dvs andet neuron, fortsætter til glatte muskelceller eller kirtelceller repræsenteret i kar eller organer. En tilsvarende omkobling sker ikke for nervetrådene (6) til tværstribet muskulatur. I det somatiske nervesystem nås effektorcellen af første neuron. I det autonome nervesystem bruges normalt to neuroner. | |
| Nogle nervetråde (2) fra lateralhornene afbrydes ikke i grænsestrengsganglier. Trådene går igennem grænsestrengen (1) og afbrydes først i “udflytterganglier” (3). Vigtigst er nn. splanchnici (2) fra grænsestrengen i cavitas thoracis. De går gennem diafragma (11) til “solarplexus” (plexus coeliacus), lige under diafragma. Trådene afbrydes i ganglier (3) (ganglia coeliaca), der er indlejret i plexet. Neuriterne fra de nye celler følger karrene til organerne (12‑19). | |
Side 380
| Alle sympatiske nervetråde begynder med udspring fra lateralhorn i medulla spinalis, dvs, alle tråde kommer fra thoracalsegmenterne og de to øverste lumbalsegmenter. | |
| De sympatiske tråde til hoved og hals løber opad gennem højre og venstre side af grænsestrengen og danner synapse med andet neuron i et ganglie i halsdelen (4) af grænsestrengen, truncus sympaticus. | |
| Truncus sympaticus omfatter i halsdelen tre ganglier (1), (3), (4), hvoraf det øverste er størst. | |
| Fra gangliecellerne i halsdelen (4) går sympatiske nervetråde til kirtler (5) på hoved og hals, kar (6) på hoved og hals, musklen der udvider pupillen (8), m. dilator pupillae, hjertet (10) og bronchierne (9). | |
| Det kan virke ejendommeligt, at der går sympatiske nervetråde til hjertet (og andre brystorganer) fra halsdelen af truncus sympaticus. Men det er logisk nok, fordi hjertets udvikling begynder opadtil på halsen lige under den primitive mundhule, og allerede på det tidspunkt vokser nerverne frem til hjertet. Når hjertet senere indtager sin endelige plads i brystkassen, trækker det nerverne fra halsen med sig. | |
| Lige så ejendommeligt virker det, at nerverne til musklen, der udvider pupillen (8), m. dilator pupillae, udspringer fra lateralhorn i rygmarvens brystdel. De løber straks til truncus sympaticus (1) i cavitas thoracis og fortsætter derefter til øverste ganglie i halsdelen (4), hvor der er synapse med andet neuron, der følger nervetråde (5) langs blodkar til musklen i øjeæblet (8). | |
| Det sympatiske system aktiveres som sagt især ved forskellige stress situationer, og den samlede virkning er, at kroppen bedre er i stand til at imødegå krisesituationer. Hjertets evne til at pumpe blod øges samtidig med, at blodkarrene i huden og de indre organer trækkes sammen, så en større del af blodet strømmer til skeletmusklerne. Øget sympatisk aktivitet bedrer også ilttilførslen, idet bronchierne udvides, og åndedrætsfrekvensen øges. | |
| Sympaticus fremmer blodtilstrømningen til hjertemuskulaturen ved at dilatere hjertets egne kar. Alle andre steder virker sympaticus karkontraherende. Det udnyttes i tandlægepraksis ved lokalbedøvelse, der sker i områder med en meget rigelig blodgennemstrømning. Sammen med bedøvelsesvæsken indsprøjtes derfor et stof med samme virkning som sympaticus for at få blodkarrene til at trække sig sammen. Derved opnås, at bedøvelsesvæsken bliver i længere tid på det sted, hvor den skal virke. | |
Side 381
| Det parasympatiske nervesystem dominerer i hvilesituationer. Det nedsætter hjertets pumpeaktivitet og blodtryk, øger peristaltikken, dvs transporten gennem mavetarmkanalen, og øger sekretionen fra kirtler fx i tilknytning til fordøjelseskanalen. | |
| De parasympatiske nervetråde fra kerner (1‑4) i hjernestammen løber perifert i hjernenerverne nr. 3, 7, 9 og 10. Langt de fleste parasympatiske tråde findes i hjernenerve nr. 10, n. vagus (4). | |
| Parasympatiske tråde i hjernenerve nr. 3, n. oculomotorius, løber til det parasympatiske ggl. ciliare (5) bag øjeæblet. Gangliet består af små multipolare nerveceller, hvor de parasympatiske tråde (17) afbrydes. Neuriterne går til øjeæblet, hvor de innerverer ringmusklen, der forsnævrer pupillen (7), m. sphincter pupillae, og akkomodationsmusklen (6), m. ciliaris, der ændrer linsens krumning. | |
Side 382
| Nogle af de parasympatiske tråde i hjernenerve nr. 7, n. facialis, løber til det parasympatiske ggl. pterygopalatinum (3), der ligger i fossa pterygopalatina lige under n. maxillaris (1). | |
| De parasympatiske tråde danner synapse med andet neuron i ggl. pterygopalatinum (8), og neuriterne fra cellerne i gangliet fordeler sig gennem grene fra n. maxillaris og n. ophthalmicus til gl. lacrimalis (9), kirtler i cavitas nasi (10) og i ganen (11). Andre parasympatiske tråde i n. facialis (2) går til ggl. submandibulare (12) i trigonum submandibulare. Trådene afbrydes i gangliet, og neuriterne fra andet neuron går til to store spytkirtler, gl. sublingualis (13) og gl. submandibularis (14). | |
| De parasympatiske nervetråde i hjernenerve nr. 9, n. glossopharyngeus, går til ggl. oticum (22). Gangliet ligger på bagsiden af n. mandibularis lige under foramen ovale (1), dvs bagved pterygoidermuren og i den allerøverste del af spatium lateropharyngeum. | |
| De parasympatiske tråde danner synapse med de små multipolare celler i gangliet (15), og neuriterne i andet neuron går til gl. parotidea (16), idet de løber sammen med n. auriculotemporalis fra n. mandibularis. | |
| Tråde fra den parasympatiske kerne (4) knyttet til hjernenerve nr. 10, n. vagus, fordeles til organer i hals, brysthule og bughule, til midt på tyktarmen ved flexura coli sinistra. N. vagus virker hæmmende på hjerte og lunger i brysthulen og fremmende på organer i bughulen, særlig på sekretion og peristaltik. N. vagus har ingen parasympatiske ganglier, men trådene danner synapse med små multipolare nerveceller, der ligger spredt i væggene af de organer (18‑26), nerven forsyner. Om nervetrådene i det parasympatiske system gælder, at de har et langt forløb, inden de danner synapse med andet neuron. Det er mest udpræget for n. vagus, hvor de parasympatiske tråde har et overordentligt langt forløb, inden de danner synapse i organvæggene. | |
Side 383
| Den anale del af tyktarmen, dvs fra flexura coli sinistra, samt bækkenorganerne får parasympatiske tråde (29) fra rygmarvens sacraldel. Trådene afbrydes i organernes vægge. | |
Side 384